The Antiquity of the Piltdown Race
The Antiquity of Man Vol. II 1925
Arthur Keith
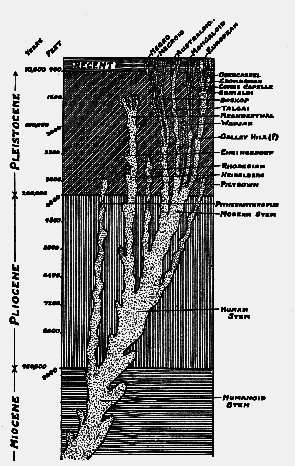
Geological tree indicating the ancestry of living and extinct races of mankind. On the left are given estimated depths of the more recent geological deposits and of the time occupied by their accumulation.
[503] CHAPTER XXVII
THE ANTIQUITY OF THE PILTDOWN RACE
A great company assembled in the rooms of the Geological Society of London on the evening of 18th December 1912, to receive the first authentic account of the discovery at Piltdown. An unknown phase in the early history of humanity was to be revealed; a revelation of that kind stirs the interest of many men, and draws them from their studies and laboratories to brave the heated atmosphere of overcrowded meeting-rooms. The various fragments of the skull had been pieced together; the missing parts had been filled in; a complete skull was thus brought before the meeting. It was quite plain to all assembled that the skull thus reconstructed by Sir A. Smith Woodward was a strange blend of man and ape. At last, it seemed, the missing form–the link which early followers of Darwin had searched for–had really been discovered. No one had ever suspected that a secret of this kind lay hid away in the Weald of Sussex. We shall attend the meeting of geologists, however, not so much to learn what kind of beings those ancient inhabitants of England were, as to ascertain their position in the scale of time–to see their place in the scheme of man's evolution. We want to hear from the lips of those who have studied the recent history of the earth, and who have discovered the sequence and the dates of more recent land changes and deposits, how long ago it is since these ancient people lived in the Weald of Sussex. We shall take, in the first place, the opinion of Sir A. Smith Woodward. In his judgment the Piltdown remains "are almost (if not absolutely) of [504] the same geological age" as the Heidelberg mandible– the oldest fragment of man's body yet found on the continent of Europe. The geological age of the Heidelberg mandible, as we have seen in a former chapter, has been fixed with some approach to accuracy. It was discovered in a deposit laid down in the ancient bed of a tributary of the Neckar near the beginning of the Pleistocene stage of the earth's history. The Cromer beds were in process of formation in East Anglia. Sir A. Smith Woodward's opinion, then, is that the Piltdown form of man was living in Southern England at an early part of the Pleistocene period, and that, at the same date, a very different kind of man was inhabiting Central Europe.
The reader may naturally break in with the question: How long ago is that? The facts which will yield an estimate of geological time certainly exist, and in the opinion of men like Rutot, Sollas, and Penck are sufficient to afford an approximate estimate–the first step towards accurate figures. We shall take the estimate of Professor Sollas 1 first. He regards the deposits which were laid down during the Pleistocene period as forming, when superimposed, a thickness or depth of 4000 feet (see frontispiece). He estimates that the formation has proceeded at the rate of a foot per century, and that therefore the collective deposits of the Pleistocene period probably have taken about four hundred thousand years to form. The estimate given by Professor Rutot is much less–one hundred and forty thousand years. Short as it is, that estimate deserves our serious consideration, for it is founded on a prolonged study of the Pleistocene formations found along the river valleys of Belgium. There is a third estimate which must also weigh with us in coming to a conclusion as regards the duration of the Pleistocene period–that of Professor Penck. 2 He has studied the changes produced by Alpine glaciers during the Pleistocene cycles of extreme cold. He is of opinion that such changes indicate for the Pleistocene [505] period a duration of at least half a million years– perhaps they may have occupied as much as a million and a half. These figures are mere provisional estimates, subject to modification as our knowledge increases. The numerous changes in climate, of elevation and depression of the land, the transformation of our animals, the elaboration of human culture, the evolution and distribution of human races, all bespeak an enormously long period of time.
To arrive at an estimate of the antiquity of the Piltdown remains, we must also allow for the time which has elapsed since the Pleistocene period ended and our present era began. There is now a general agreement that about ten thousand years would cover this recent period. When, therefore, Sir A. Smith Woodward assigns the Piltdown remains to an early phase of the Pleistocene epoch, we may, in the present state of our knowledge, suppose him to refer the Piltdown race to a time which passed, certainly one hundred thousand years ago, perhaps as much as five times this sum. There is a growing tendency amongst geologists to shorten the length of the Pleistocene period. Beyond any question, the Piltdown skull represents the most ancient human remains yet found in England.
Having thus attempted to give Sir A. Smith Woodward's opinion of the antiquity of the Piltdown remains m terms of years, we must again return to the meeting of geologists and take up the narrative there. The discoverer of the remains, Mr Charles Dawson, 3 said "he was quite prepared, from an anthropological point of view, to accept an earlier date for the origin of the human remains, and Dr Woodward and he had perhaps erred on the side of caution in placing the date as early Pleistocene." In Mr Dawson's opinion, then, it is possible that the Piltdown race may belong to the period preceding the Pleistocene–the Pliocene. Sir William Boyd Dawkins said "he agreed with the authors of the paper that the deposit containing the human remains belonged to the [506] Pleistocene age, and that the Pliocene mammalia in it– Mastodon arverensis and the rest–had been derived from a Pliocene stratum formerly existing in that area." This opinion, coming from one who has the right to speak with authority, must evoke surprise. When he found the remains of the same species of Mastodon in the Doveholes cave in Derbyshire in 1903, unaccompanied by human remains, Sir William unhesitatingly assigned the contents of that cave to the Pliocene period; but when the same remains are found in Sussex, accompanied by human remains, the deposit, in his opinion, should be referred to a much later date. In Sir William's opinion man is an evolutionary product of the Pleistocene period, and first reached Britain about the middle of this epoch. Much more guarded opinions were given by Mr Clement Reid and by Mr A. S. Kennard, who have made a special study of the later deposits in the south of England. "It was impossible," said Mr Reid, "to speak with confidence, but the whole evidence suggested that the Piltdown deposit and the plateau on which it rests are not preglacial or even early Pleistocene; they belong to a period long after the first cold period had passed away, but they occur at the very base of the great implement-bearing succession of Palæolithic deposits in the south-east of England." Mr Kennard regarded the Piltdown gravel as being of the same age as the 100-foot terrace of the Thames valley, which, as we have already seen, is made up of strata belonging to various stages of the Pleistocene epoch. Mr Kennard's opinion is of the greatest interest, because it was from the 100-foot terrace of the valley of the lower Thames, at a depth of 8 feet, in 1888, that a human skeleton was found at Galley Hill. If Mr Kennard is right, the Galley Hill man, who was of the same type as modern man, must have been almost contemporaneous with the very primitive human being which Sir A. Smith Woodward had reconstructed from the Piltdown fragments. Mr E. T. Newton, who, in 1896, brought the Galley Hill discovery before the same [507] Society as now discussed the Piltdown find, was also present. It must have puzzled him to explain why the audience, which in 1896 refused to accept the Galley Hill discovery, because the remains were those of a being framed much as we moderns are, should extend so ready an acceptance to the very simian form of man which had been raised from the Piltdown fragments. Here we are concerned only with the opinion Mr Newton formed of the antiquity of the Piltdown remains. To him, "the highly mineralised condition of the specimens seemed to point to their being of Pliocene age rather than Pleistocene."
We must also note the opinion of the leading authority of France–Professor Marcellin Boule. He agrees with Mr Kennard in regarding the Piltdown gravels as of the same age as the 100-foot terrace, and while in his text he assigns Piltdown man to the time of Chellean culture, yet in the table, where the succession of Pleistocene men is shown, Piltdown man is placed later than Heidelberg man, being made to coincide with the earliest age of Acheulean culture. 4
The writer is a student of the human body, and is therefore not in a position to offer any conclusive evidence which will help to settle whether the Piltdown man was Pleistocene or Pliocene. Yet there is one point which must weigh with those who seek to place this newly discovered human form in its proper place in the scale of time. The lower jaw, especially in the region of the chin, is marked by certain characters which separate it sharply from the corresponding part of all human mandibles and link it closely with the jaw of apes. Even in the Heidelberg mandible, which belongs to the early Pleistocene age, the essential features of humanity are already evident. In the Piltdown mandible the conformation is that of the ape; a simian stage is still preserved. The Heidelberg mandible shows that the human contour of the chin had already appeared at the beginning of the Pleistocene, but a change of this kind has not become [508]
manifest in the Piltdown mandible. This feature suggests that Piltdown man represents, as the animal remains accompanying him suggest, a Pliocene form. I am of opinion that future discoveries will prove that the remains found at Piltdown represent the first trace yet found in Europe of Pliocene man.The reader may feel by the time he has reached this point that enough has been said about the time at which the Piltdown man lived. Probably he is already wearied with the clash of expert opinion. Yet the differences are perhaps not so great as they appear. It will be remembered that Mr Lewis Abbott, who has given as much time as anyone to master the later geological history of the Weald, expressed the decisive opinion that in the Piltdown gravel two ages are represented. The lower or bottom stratum, which contained the Pliocene remains and human bones, is, in Mr Abbott's opinion, Pliocene in date; the upper levels, in which the rude Palæolithic implements lay, have been disturbed at a later time, and are to be regarded as Pleistocene in age. If Mr Abbott is right and a survey of the full evidence favours his inference, then the divergence of opinion is explicable: those who maintain that the Piltdown gravel is Pleistocene are right, and so are those who regard it as Pliocene. Indeed, in a subsequent communication Mr Dawson wrote of the "dark" or Eoanthropus stratum as follows:
5 "We cannot resist the conclusion that the third or 'dark' bed is, in the main, composed of Pliocene drift, probably reconstructed in the Pleistocene epoch. . . . Putting aside the human remains and those of the beaver, the mammalian remains all point to a characteristic fauna of Pliocene age; and, though all are portions of hard teeth, they are rolled and broken. The human remains, on the other hand, although of much softer material, are not rolled, and the remains of the beaver are in a similar condition."So far we seem to have gone a long way merely to reach the conclusion that the Piltdown man is possibly [509] of Pliocene age. To assign even the remains of man to the Pliocene period carries but a shadowy significance to most of us. If, however, we again visit Piltdown and survey the changes which have occurred in the Weald since fossil man was living there, we obtain some insight into his great antiquity. Mr Dawson discovered and delimited the remains of a great sheet of gravel which, in former times, covered the Piltdown plateau. The sheet apparently extended (see fig. 167) for about twelve miles in one direction and eight in another, and must have formed a great deposit for such a small stream as the predecessor of the Sussex Ouse to lay down. Only some patches and pockets of the original deposit have come down to us. The plateau is now 120 feet above the level of the sea, but it is probable that it lay almost at sea-level when this great sheet was being deposited. The formation of so extensive a gravel bed must have occupied a long space of time, for the oldest or bottom layer is apparently Pliocene in date the upper or disturbed layer is much later, probably middle Pleistocene in age. Over this wide Pliocene plain the ancient Ouse had meandered, shifting its bed from time to time and laying down gravel, sand, and fossil remains, gathered on the higher lands of the Weald. The present Ouse has cut a valley, 80 feet deep, in the plateau. This valley has been excavated since the time the more recent gravel beds were laid down on the plateau. How often the valley has been re-excavated, as the land rose; how often it has been filled up, as the land sank, we cannot as yet tell; certainly the valley of the Ouse rose and fell with that of the Thames. But it is clear that there have been many variations in the level of the land since the gravel was first laid down on the Piltdown plateau. The human remains lay in the most ancient gravel deposit. Since the Piltdown man lived, then, the great expanse of gravel, measuring nearly one hundred square miles, has been laid down and a valley, at least 80 feet deep, has been slowly eroded by a comparatively small stream. As the first gravel was being laid down the culture of man was represented by [510] flints worked in a pre-Chellean style and surprising pieces of bone work. And long before Piltdown days there had been the men who shaped eoliths. As things are to-day, man's culture is represented by the wireless messages and aeroplanes which cross the Weald, and the great steamers passing down the Channel, and the rural homes and country houses which everywhere meet the eye.
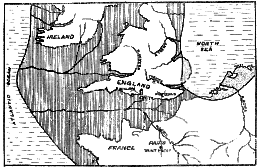
Fig. 171.–Map of South England and North France, to show the course and tributaries of the ancient channel river. (After Boyd Dawkins.)
No one suspected, until Mr Dawson made the discovery, that deposits of a Pliocene or early Pleistocene date occurred in the Weald of Sussex. It is not likely that Piltdown is the only site at which such deposits occur in the region of the English Channel. If one looks at a map representing the bounds of England during the great land elevation in late Pliocene times–the date at which Piltdown man or his ancestors may have lived–it will be seen (fig. 171) that in place of the English Channel there is a great river which is joined by all the streams issuing from the southern area of the Weald. It will be seen, too, that the Somme and the Seine also lie within the watershed of the great channel river. Now on a tributary of the Seine is situated St Prest, [511]
near which there is, as M. Rutot has indicated, a deposit very similar in nature and in age to that at Piltdown. We have already seen that the deeper strata of gravel at St Prest contain remains of Pliocene animals and have always been regarded as of a Pliocene age (see fig. 114, p. 317). Over the Pliocene beds are others of a later or Pleistocene age, just as at Piltdown. So long ago as 1863, M. Desnoyers 6 recognised that many of the fossil bones of animals existing in the Pliocene period, and found in the deepest and oldest deposits of St Prest, showed definite evidence of having been worked by man. More striking still, the land on which these deposits occur is 80 feet above the adjoining tributary of the Seine, the Eure–the same height as the Piltdown deposits lie above the Ouse. The Pliocene age of the St Prest deposits has never been called in question.There is a further striking similarity between the deeper deposits at St Prest and the dark bed at Piltdown. Mention has been made of the eoliths which occurred with the human remains in the dark bed. Shaped flints of a corresponding type also occur in the Pliocene beds at St Prest. They were discovered and described by a geologist, M. Bourgeois,
7 in 1867, four years after M. Desnoyers recognised the human markings on the fossil bones. In M. Rutot's opinion the St Prest implements are of a later and more highly evolved type than the Kentish eoliths. The discoveries made at St Prest fifty years ago have a very direct bearing on the problem of the age of the Piltdown remains.The evidence of another Pliocene deposit may be cited here. In the south of England, about one hundred miles to the west of Piltdown, but still within the watershed of the old Channel river, there occurs another trace of the Pliocene period which is of the greatest importance to the student of man's evolution. This trace occurs at Dewlish, a small village in the chalky [512] uplands of Dorset (fig. 171). Near the village of Dewlish the chalk plateau, about 300 feet above sea-level, ends in a sharp bank or escarpment, about 100 feet in height, similar to the chalk brim of the Weald. On this plateau, near Dewlish, there was discovered by accident a deep fissure in the chalk which was filled with layers of sand and gravel. This fissure or trench was investigated by the Rev. O. Fisher, and has been described by him in two communications to the Geological Society of London.
8 It was found to be over 100 feet in length, with one end open on the free face of the escarpment. Its depth was found to be over 12 feet. It was narrow and filled in with sand and gravel, which do not now occur on the surface of the plateau at Dewlish. In the gravel were found remains of elephants of the ancient kind found at St Prest and at Cromer–Elephas meridionalis–which is accepted as a true representative of the Pliocene period. In the gravel also occurred certain flints which were regarded by Mr Grist 9 as similar to the eoliths of the Kentish plateau. Mr Clement Reid inspected the trench –the only one of the kind known–and formed the opinion that it did not represent any cleft or fault produced by natural agencies. No stream could have produced such a trench; there is no stream now on the plateau. Mr Fisher could only account for it on the supposition that it was dug by the hands of man, and was designed, like similar trenches at the present day, as an elephant trap. If Mr Fisher's inference is right, and no other satisfactory explanation has been offered, we have the startling revelation that in the Pliocene period mankind had already reached an advanced stage in his evolution. And those opinions were formed and expressed years before Mr Dawson had found a bone implement at Piltdown, apparently worked from the thigh bone of [513] the kind of elephant whose fossil remains occur in the trench at Dewlish.We have already mentioned the sub-Crag implements discovered by Mr Reid Moir (p. 308). They indicate the existence of human beings towards the middle of the Pliocene period. The Kentish eoliths are more ancient than the sub-Crag implements.
10 When, therefore, we take into consideration these facts, and the similarity between the Piltdown and St Prest deposits, we are persuaded that Mr Dawson and Sir A. Smith Woodward were ultra-cautious in assigning a Pleistocene date to the human remains found at Piltdown. All the evidence seems to point to a Pliocene age–to an age at least as old as the Cromer forest bed. Hence the importance of their discovery, for although the handiwork of Pliocene man has been recognised for a considerable number of years, the man himself was unknown until Mr Dawson brought the Piltdown type into the light of day.______________________________________________________________________________
1 Nature, 1900, vol. lxii. p. 481.
2 See A. Penck, Zeitschrift für Ethnologie, 1908, vol. xl. p. 390.
3 Quart. Journ. Geol. Soc., 1913, vol. lxix. p. 151.
4 Fossil Men, translated by Jessie and James Ritchie, 1923.
5 Quart. Journ .Geol. Soc., 1914, vol. lxx. p. 85.
6 M. J. Desnoyers, Compt. Rend., 1863, vol. lvi. p. 1073. See also references given on p. 317.
7 See Paléontologie humaine, by E. T.. Hamy, Paris, 1870, p. 98.
8 Quart. Journ .Geol. Soc., 1888, vol. xliv. p. 819; 1905, vol. lxi. p. 35. Since the above was written, the Dorset Field Club has reopened the trench. The evidence favours natural–not a human–formation.
9 C, J. Grist, "Some Eoliths from Dewlish, and the Question of Origin," Journ. Roy. Anthro. Inst., 1910, vol. xl, p. 192
10 J. Reid Moir, Nature, 1924, vol. cxiii. p. 461.
[514]
CHAPTER XXVIIIEOANTHROPUS DAWSONI
Having thus settled, so far as the evidence will permit, the approximate position of the Piltdown man in the scale of time–and beyond question he represents the earliest specimen of true humanity yet discovered–we now proceed to see what sort of being he was. The truth is that we have to discover his characters from fragments of the skull, for no other part was found. The form of his limbs and body are matters of inference. The reader will quickly realise the number and size of the actual parts of the skull which were found, by examining fig. 172; the missing parts are indicated by stippled lines. The bone which forms the forehead–the frontal bone–is only partly present. Fortunately, the region which forms the upper margin of the left orbit has been preserved in its outer part, so that we can form a definite opinion as regards the supra-orbital ridges. These are not formed as they are in the chimpanzee, gorilla, Rhodesian and Neanderthal man, but are more like the conformation seen in modern human races. A great part of the left side of the frontal bone has been recovered; the right side is wholly missing, but we know that the right and left sides of the frontal bone are nearly symmetrical, so we can reconstruct the greater part of the forehead with some degree of assurance–all except the middle part lying over the root of the nose. This initial blank in our knowledge was also made good, for the fragment of the frontal bone, found by Mr Dawson in 1915, gave us the contour of parts above the root of the [515]
nose and showed that in the modelling of his forehead Piltdown man resembled living man.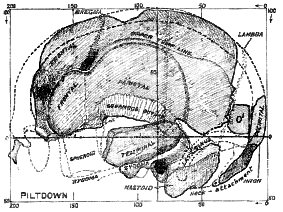
Fig. 172.–Fragment of the Piltdown skull placed in position and represented in profile.
Taking the bones which form the roof of the skull just behind the frontal bone, we see that practically the whole of the parietal bone is present on the left side. It forms the roof and side of a great part of the brain chamber and is therefore of the utmost importance. We want especially to know about the brain, for this is the master organ of the human body. It will be noted that even on
the left side certain fragments of the parietal bone are missing along its upper margin, hinder border, and at its lower angle behind. The dimensions of these missing parts can be estimated with accuracy. The parietal bone of the opposite or right side is also represented. Only about two-thirds of that bone is present, and, unfortunately, the part which is missing is the upper area which reaches up to the middle line on the roof of the skull, where it should come into contact with the opposite or left parietal bone. The actual part of the right [516] parietal bone recovered is indicated in fig. 173, where it has been superimposed over the corresponding area of the left side.
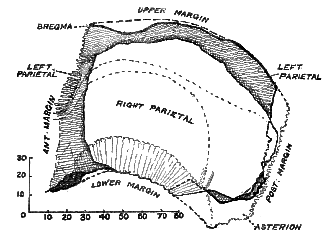
Fig. 123.–A reverse or mirror-image of the fragmentary right parietal bone of the Piltdown skull superimposed on the corresponding points of the bone of the left side, to show the extent of the parts missing from each.
Coming now to the hinder or occipital end of the skull, we find the bone of this region represented by a considerable fragment. The part of the occipital bone is best seen in a hinder view of the skull such as is shown in fig. 174. To the lower part of the occipital bone the neck is fixed; part of the region for the fixation of the neck is represented in the fragment found (fig. 172). At the lower end of the fragment is seen the hinder margin of the foremen magnum, by which the spinal cord makes its exit from the brain cavity to enter the spinal column. The tabular part of the occipital bone, which rises up from the neck to form the projecting hinder [517
] part of the head, has also an important representation. By great good fortune, Sir A. Smith Woodward recovered a most essential fragment of the right half–the fragment which gives us an indication of the width of the occipital bone, almost up to its articulation or point of contact with the right parietal bone (see figs. I72 and 174, O'). On the occipital bone the ridge which marks the middle line of the head and of the neck is preserved (fig. 174). We therefore know the width of the right half of the occipital bone, and we may be certain the left half was almost exactly of the same size. Hence in the drawing (fig 172) of the recovered fragments the right half of the occipital bone is represented as if it were on the left side, in place of the right.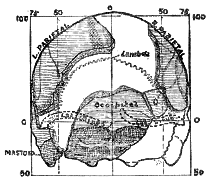
Fig. 174.–The fragments of the Piltdown skull viewed from behind. The fragment of the occipital bone, marked O', is that found by Sir A. Smith Woodward Above it is seen the hole made by the blow of the pick which detached this fragment from the parietal. I have left a narrow triangular gap between this fragment and the main occipital bone because an examination of the original specimen led me to think a splinter is missing. Compare with fig 190, B, where a reconstruction is given with fragment O', in the position assigned to it by Sir A Smith Woodward.
The frontal, parietal, and occipital bones form the front, roof, and hinder part of the brain chamber. The [518] temporal bone, which helps to complete the lateral wall of the chamber, also forms part of the floor or base of the cranial cavity. Almost the whole of this bone was found of the left side (fig. 172). A fragment is missing, but sufficient of its upper border is preserved to give us its true relationship to the parietal bone. As may be seen in fig. 172, a mutual point of contact is present on the opposing margins of the temporal and parietal bones. It will thus be realised that all the essential components of the brain chamber are present; there are sufficient parts to form almost a complete half of the brain chamber. In the case of the skull a half is almost as good as a whole, for the left and right halves of every head are approximately similar.
All of these fragments of the Piltdown skull are, at first sight, very similar to the same parts of a modern human skull except as regards their thickness. The Piltdown bones are surprisingly thick. The farm-labourer who first saw the skull described it as a coco-nut. In most modern heads the thickness of the bones forming the brain chamber varies from 4 to 6 mm. (1/6 to 1/4 of an inch); in native African races, and occasionally in Europeans, the thickness may amount to 8 or even 10 mm., but in no normal modern skull are all the bones so uniformly thick as in this recently discovered specimen. As already pointed out, the ancient skulls found at Galley Hill, Clichy, and Olmo are thick, but not to the extent seen in the Piltdown fragments. Thickness is also characteristic of most Neanderthal skulls. In the Piltdown cranium the frontal, parietal, occipital, and temporal bones vary in thickness from 8 to 12 mm., the average all over being about 7, of an inch. The bone is naturally formed; there can be no question of disease. The late Professor Shattock definitely settled this point.
l There can be no doubt that sufficient of the skull has been recovered to provide us with the means of reaching a just and certain conclusion as regards the size [519] and shape of the part which contains the brain. Very few ancient skulls are so well represented as that of Piltdown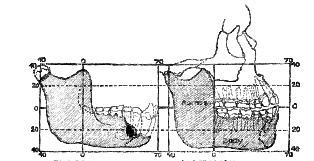
PILTDOWN AUSTRALIAN
Fig. 175–The Piltdown mandible, as seen in true profile, compared with a corresponding view of the mandible of an Australian native. The missing teeth and parts of the Piltdown mandible are indicated by stippled lines.
The discovery of almost a complete half–the right–of the lower jaw or mandible by Mr Dawson is a most fortunate circumstance (fig. 175). It lay in the iron-stained cemented stratum with unworn eoliths and a fragment of the tooth of an early Pliocene form of elephant. The importance of the mandible is at once apparent; it provides us with the skeletal outline of the face of this ancient form of man. Each half of a human mandible consists of two distinct parts: (1) a horizontal part or body (fig. 175), which carries the teeth and forms the lower part of the outline of the face–from the angle of the jaw below the ear to the chin; (2) a vertical part, which ascends from the angle to terminate in an articular knob or condyle. The socket–the glenoid cavity–for the articular knob is situated on that part of the base of the skull which is formed by the temporal bone immediately in front of [520]
the ear. We have already seen that the temporal bone of the left side was recovered, revealing the characters of the articular cavity, which is shaped exactly as in modern races, and in this respect quite unlike the same joint in an ape's skull. In the Piltdown skull we have thus the greater part of one-half of the mandible of the right side; on the left side the articular socket for the jaw is present; by transposing or reversing the right half of the mandible to take the place of the left half it will be seen that we obtain a representation of the skeletal outline of one-half of the Piltdown head (see fig. 252, p. 69I)Unfortunately, a fragment is missing from each part of the right half of the mandible (fig. 175). All of the ascending part is present; the areas for the attachment of the chewing muscles are intact, save the articular knob itself. This is no great loss, because the shape of the socket in which it plied is known to us, and as that socket, although large, is shaped as in present-day man, we may presume the articular knob or condyle of the jaw also had a modern human form. The part missing in the body or horizontal part of the jaw leaving some room for speculation. That region of its upper border which lies hidden behind the lower lip in life, and carries the sockets of five teeth–the two right incisors, the right canine, and two right premolar teeth, has been broken away. The part which bears the three molar teeth is present; the first and second molar teeth fill their sockets, but the third or wisdom tooth had dropped out after death. The deficiency thus affects the upper or alveolar border of the jaw in front; the whole of the lower border of the body of the mandible is present. As the body approaches the region of the chin (see fig. 238) it becomes modelled in a form which is peculiar to apes.
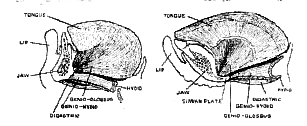
Early in the summer of 1912, when Sir A. Smith Woodward commenced his examination of the Piltdown fragments, he realised that the peculiar and characteristic features of this ancient form of man were centred in the [521] region of the chin. Such features had never been found nor seen in any mandible or skull to which the term human could be applied. The peculiar characters of the chin, which arrested his attention, can best be realised by a reference to such illustrations as are shown in figs. 176 and 177. In fig. 177 is represented the arrangement of parts seen in the chin region of a young chimpanzee–about four years of age, just before the milk teeth are shed.
MAN CHIMPANZEE
Fig. 176.–Section of the human tongue, chin, Fig. 177.–A corresponding
lower jaw, and lip made along the middle line, section of the same region of a
to show the origin of muscles from the region of young chimpanzee.
the chin or symphysis.
When a section is made so as to separate the lower jaw and the tongue into right and left halves, it is seen that the jaw in the region of the symphysis–the line of fusion of the right and left halves of the jaw–is composed of two parts, an upper part to bear the incisor teeth, and a lower part which is for the attachment of muscles. It is the lower or muscular part which principally concerns us. There is no projection of the anterior surface at the lower border of the symphysis to represent a chin in the chimpanzee; the anterior or labial surface of the jaw slopes downwards and backwards to a chinless lower border. On the hinder surface of the symphyseal region–the surface directed towards the tongue–there is seen a deep pit, almost large enough to take the tip of the little [522]
finger. From the interior of the genial pit arise the two chief muscles of the tongue–the genio-glossus muscles. The lower margin of the pit is formed by a plate of bone–the simian plate–which unites the lower borders of the right and left halves of the jaw (fig. 177). It is a strengthening plate. From the posterior margin of the plate two pairs of muscles take their origin–the genio-hyoid muscle, which draws the larynx forward during the act of swallowing, and the digastric muscle, which depresses the front part of the jaw, and thus assists in opening the mouth. Such is the conformation of the symphyseal or chin region of the lower jaw in apes (see fig. 177).When a corresponding section is made of the symphyseal region of a human lower jaw, a very different conformation is seen (fig. 176). There are the same two parts exposed in the section–the upper dental part, which carries the incisor teeth, and a lower part for the attachment of muscles. The dental part, compared with that of an ape, is small and vertical in direction. The human teeth being relatively small, the dental part of the jaw has retreated backwards in the mouth. The muscular part, on the other hand, has been advanced; there is now a prominent, or, at least, a well-marked chin. The genial pit and simian shelf seen in the ape's jaw are absent. In place of rising from a pit, the main muscles of the tongue–the right and left genio-glossus–arise from an elevation of bone bearing two tubercles. The genio-hyoid, in place of rising from the upper margin of the shelf of bone springs from the lower part of the elevation which has filled up and replaced the pit, while the digastric muscles are attached to the lower border of the jaw just behind and below the chin.
It will thus be seen that the simian and human mandibles differ markedly in the region of the symphysis or chin. The meaning of this difference will be discussed in a later chapter (p. 659). As to the type to which the Piltdown jaw belongs there can be no doubt; both the genial pit and the simian plate are present. These are ape-like [523
] features. Sir A. Smith Woodward recognised them as such, and in his work of reconstructing the original form of the skull the presence of these simian features exercised a dominating influence. Hence, when he came to replace the missing parts of the jaws and the incisor and canine teeth, he followed simian rather than human lines. The teeth of man form a uniform series; there is no break nor diastema in front of, or behind, the canine teeth; the canine tooth does not project prominently beyond its fellows.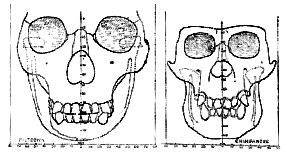
Fig. 178.–The muzzle and front teeth of the Fig. 179.–Similar view of
Piltdown skull as originally reconstructed by Sir A. the same part of a male
Smith Woodward. chimpanzee.
From the ape-like features of the chin it was inferred that projecting simian canine teeth must have been a characteristic of the Piltdown form of man. A massive canine tooth was therefore modelled; not very projecting (see figs. 178, 180), thick and rather stumpy, its longest or front-to-back diameter being 14.5 mm., its side-to-side, 10 mm. The diameters of such a tooth are equal to those of the canine of a male chimpanzee, and far beyond the limits of the largest human canines known As in the chimpanzee, a break or interval in the dental series was left both in front of and behind the canine. The width and conformation given to the front or incisor teeth were those of the chimpanzee; when the Piltdown [524] lips parted one would have seen the ferocious dental array of the anthropoid (figs. 179, 181). As regards the rest of the teeth–the two premolar and three molar–man's dentition rather than the chimpanzee's was copied. The first and second molar teeth were found actually in the jaw; they were not larger than the corresponding teeth of certain modern races, and are distinctly human in pattern–at least more human than anthropoid. The premolars had to be made much larger than in human jaws, but they were given a human not a simian form. Thus in the first reconstruction of the Piltdown skull there appeared to be a mixture of dentitions. In front the teeth were simian; behind they were human.
We are dealing at present with the parts which were actually found when the discovery was first announced. In the autumn of 1913 a canine tooth was discovered which must be assigned to the Piltdown mandible. It was, as Sir A. Smith Woodward had anticipated, essentially simian in form–it was more like that of a female chimpanzee or gorilla than that of a human being. By the time this tooth was discovered I had come to the conclusion, for reasons which will be given in another chapter. that a massive human and not a projecting and simian canine had really been present. In that I was mistaken. but as regards the actual dimensions of the tooth my estimate was approximately right. I allowed 10 mm. for the longest (front-to-back) diameter; it proved to be 11 mm. Sir A. Smith Woodward had represented it as 14.5 mm. In the chief point, however, Sir A. Smith Woodward was right; the simian chin was correlated with a simian canine tooth.
As will be seen from figs. 178 and 179, Sir A. Smith Woodward made the muzzle and front teeth of Eoanthropus wider and more massive than in the chimpanzee. He made the region of the chin and symphysis–the anterior line of fusion of the two halves of the mandible particularly strong (fig. 180). The symphyseal areas of union or fusion between the two sides of the mandible are stippled in figs. 180 and 181. In the Piltdown man[525
]dible this area, as restored by Sir A. Smith Woodward, is even greater than in the chimpanzee. While the teeth implanted in the front part of the mandible and the symphyseal region are truly simian, the hinder part of the mandible, the molar teeth, and also the ascending branch or ramus, are, to my eye, entirely human. We have thus in this newly discovered form of man a remarkable mixture of simian and human characters.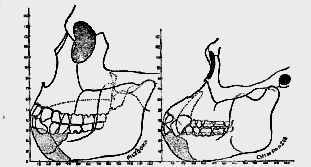
Fig. 180.–Drawing of the jaws, in profile, from Fig. 181.–A drawing of
Sir A. Smith Woodward's original model of the the same part of a male
Piltdown skull. chimpanzee.
One other feature may be pointed out here. It will be observed in fig. I75 that the greater part of the last molar or wisdom tooth lies behind the anterior margin of the ascending ramus of the jaw, being thus hid from view. In the Australian jaw (shown in the same figure), as is usually the case in man and apes, this tooth lies wholly in front of the ramus and is freely exposed. The Piltdown ascending ramus is remarkably wide (44 mm.), and its width is evidently due to a forward extension of its anterior border. On the anterior border is inserted the temporal muscle, the chief agent in biting or in suddenly shutting the lower jaw. Such a forward extension of the ramus must give the temporal muscles greater power [526
] and purchase over the front part of the jaws and canine.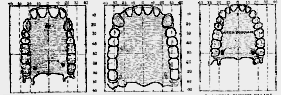
Fig. 182.–A comparison of the palatal areas in a female chimpanzee (A), in the Piltdown specimen as reconstructed in the original mould (B), and in a modern Englishman (C).
It is necessary to examine in more detail the original reconstruction of the apparatus of mastication in the Piltdown man. Our estimate of the position of any newly discovered form of human being turns largely on the relationship between his alimentary and cerebral systems. It seems fairly certain that the tendency in human evolution is to increase the work of the brain and diminish the work of the stomach. An increase of brain power has made the task of our digestive system easier– at least those parts which are concerned in mastication. We therefore need some means of indicating–even if the method adopted be but a rough approximation to the truth–the relative development of cerebral and alimentary powers in any newly discovered form of human being. We may take a concrete example. In fig. 182 is represented the palate of a female chimpanzee; the teeth are set round the circumference of the palate. Within the semicircle of teeth lies the tongue, but the whole area of the palate, all that lies inside the outer margins of the teeth, is directly concerned in mastication, and the total area of the palate may therefore be accepted as the degree to which the apparatus of mastication has been developed. We suppose that a large palate means a crude and not a richly nutritious diet. Now, in this female chimpanzee the area of the palate is 36.5 cm.
2; [527] the brain measured 320 c.c.; there was 1 cm.2 of palate to 8.7 c.c. of brain; this represents a common palato-cerebral ratio amongst man's nearest allies–the anthropoid apes. In modern Englishmen–my estimate is founded on an accurate investigation of twenty-two medical students–the average palatal area is 26.6 cm.2; the cerebral development, 1500 c.c.; the palato-cerebral ratio was therefore 1: 56.3, in place of 1: 8.7 as in anthropoids. 2 We therefore turn with some interest to see what ratio may hold in this newly discovered form of man. Sir A. Smith Woodward has reconstructed the palate. Accurate indications as to its shape were available as soon as he had obtained the form of the lower jaw and teeth, for the upper and lower jaws must fit and the teeth correspond in all higher animals, according to certain definite laws. Now the area of the palate thus reconstructed (see fig. 182, B) is 53.20 cm.2–larger than in the female chimpanzee; it is the size of palate seen in the adult male chimpanzee. In the male adult orang and gorilla the palatal area may reach 70 cm.2. Sir A. Smith Woodward at first estimated that the brain capacity of Eoanthropus was about 1070 c.c. In this new form of man the palato-cerebral ratio is therefore about 1:20. This ratio holds an intermediate position between that of the chimpanzee (1:8) and that of modern Englishmen (1:56). We appear, therefore, to be dealing with a very primitive form of man–one which, so far as concerns its development of palate and of brain, supplies us with a stage half-way between ape and man. In modern native races the palato-cerebral ratio may be as low as 1:36.7. In the Gibraltar skull, as we have seen, 3 the ratio is 1:38. In Rhodesian man this ratio is 1:31.7; in La Chapelle man, 1:41.8.We have already noted that the front teeth of this new form of man and the region of the chin are essentially [528] ape-like–quite different from those of any known form of human being. The humanity of this being, however, becomes more and more apparent as the mandible is followed backwards and upwards to its socket at the base of the skull. The socket is robust and massive, but its conformation is absolutely that seen in the more primitive of modern human races. When Sir A. Smith Woodward came to fit the fragments of the skull together, he found that the parts were human in form and must be fitted together as in modern human skulls. The shape which the reconstructed head of Eoanthropus took in his hands is shown in a series of figures (figs. 183, 185, and 188). Students of anatomy will at once recognise the peculiar features of this newly discovered type of man; but in order that those who have not made a special study of man's body may also have a standard for comparison, corresponding drawings of the skull of a modern Englishman are reproduced for comparison. The Englishman chosen has a brain capacity of 1425 c.c.–60 less than the average amount. The walls of his brain case have an average thickness of 6 mm., whereas in Eoanthropus the thickness is quite 10 mm. along the vault. Both skulls have been poised on the same horizontal plane–one which approximately indicates the lower limits of the cerebrum. The higher or cerebral part of the brain lies above the line on which the skulls are oriented.
4 In the modern head (fig. 189) the roof of the skull almost reaches the 100-mm. line; in Eoanthropus it passes slightly above the 90-mm. line. When we allow for the great thickness of the skull, it is plain that the brain of Eoanthropus will fall–as regards height–about 15 mm. (2/5 inch) short of the modern English brain. As regards the total length of the skull, the ancient and modern man are much alike–the maximum length in each case being about 190 mm.It is clear from these diagrams that the face of Eoan[529]thropus is much more massive than that of modern man. As regards details of head conformation they have much in common. In both a prominent mastoid process lies behind the ear-hole; it is shaped in the Piltdown man as in modern human races.
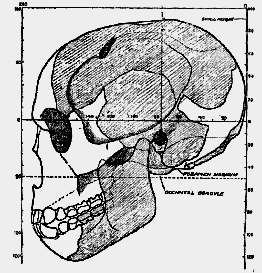
Fig. 183.–Profile drawing of Sir. A. Woodward's first reconstruction of the skull of Eoanthropus (half natural size). The parts present are shaded; the missing parts are left blank.
As the mastoid is one of the structures by means of which the muscles of the neck move and balance the head, we must infer, from the fact that this process is present in its modern shape in Eoanthropus, that the head in that ancient type of man was carried and balanced just as it now is in us. The eye-socket of Eoanthropus will be seen to be set obliquely; when the skull is viewed in true profile, more of the orbit is then to be seen than in modern human races, and much [530]
more than in anthropoid apes. Above the outer angle of the orbit it will be seen that the temporal line or crest–from which a muscle of mastication arises–ascends steeply on the frontal bone, whereas in all modern human skulls the curvature is less acute and its backward trend more marked (see figs. 183, 184). The muscles of mastication evidently did not work exactly as in modern man, or why this difference in the conformation of the temporal crest?
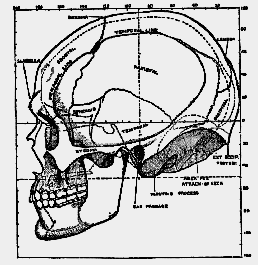
Fig. 184.–Profile drawing of the skull of a modern Englishman with a cranial capacity of 1425 C.C. (half natural size).
When a full-face drawing of the skull of Eoanthropus is compared with that of a modern skull (figs. 185, 186) a number of differences become apparent. In width, both at the base of the skull and across the cheek bones, Eoanthropus is considerably the greater. The usual width [531
] of the modern skull at its base is 132 mm.; in Eoanthropus the width here measures 150 mm. The chief difference, however, lies in the filling of the brain case. In the modern skull the sides are nearly vertical, with a slight outward bulge half- way to the vertex; in Eoanthropus the cranial cavity is so imperfectly filled that the sides lean inwards and gradually fade into a contracted crown or roof.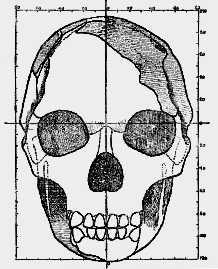
Fig. 185.–Face view of the skull of Eoanthropus as reconstructed by Sir A. Smith Woodward (half natural size). The parts shaded are those actually found.
In the modern skull, as seen from the front, the roof is usually dome-shaped. The forehead of Eoanthropus gives the impression of being contracted and ape-like; its lateral borders, formed by the lines for attachment of the temporal muscle, approximate as they ascend. In anthropoids these lines may actually meet on [532] the vertex of the skull and form a median crest. In the modern human forehead the temporal lines become wider apart, and the brow broadens towards the crown or dome of the head (fig. 176). In Eoanthropus, then, we see an ill-filled head with sides which slope inwards. In a general sense we may construe such characters as simian in nature.
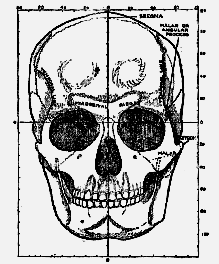
Fig. 186.–Face view of a modern human skull for comparison with fig. 185 (half natural size).
To complete this cursory review of the skull of Eoanthropus as it originally left the hands of its reconstructor, I add two further figures. In fig. 187 the modern skull is viewed from above; in fig. 188 the same view is reproduced of the skull of Eoanthropus. In the modern skull the arrangement of bones is simple. The frontal bone, forming the forehead, is joined behind to the [533]
right and left parietal bones at the coronal suture. At the posterior end of the vertical view only the upper part of the occipital bone is seen; it becomes joined to the parietal bones at the lambdoid suture (fig. 187). Along the middle line, between the right and left parietals, passes the sagittal suture. At some distance to either side of the sagittal suture is seen the upper limit of attachment of the temporal muscles–the temporal lines (fig. 187).
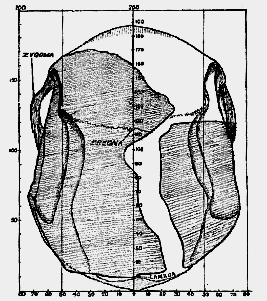
Fig. 187.–View of a modern skull from above, showing the bones and sutures of the vault (2/5 natural size). The plane of orientation is that used in fig. 184.
Part of the bony scaffolding for the support of the face and jaws is seen on the forehead–the supraorbital ridges. The cheek bars or zygomatic arches are just apparent; they pass on each side of the head in front of the ear, to end in the cheeks and thus strengthen and support the face. When we turn to the same view of the skull of Eoanthropus (fig. 188) we see representatives of the same parts, but their clear interpretation is [534] not an easy task because of the great deficiency in the forehead and along the whole length of the roof. The zygomatic arches, it will be observed, project far beyond the sides of the skull, as in anthropoid apes.
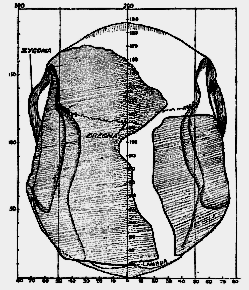
Fig. 188.–View of the skull of Eoanthropus from above, oriented on the plane shown in fig. 183. The parts shaded are those which were actually found (half natural size).
There is no definitely moulded supra-orbital ridge shown; the forehead is given a wedge shape. The temporal lines are clearly visible, both upper and lower. Only the upper of these lines are represented in the drawing of the normal skull (fig. 187). As regards general mass it is clear that the skull of Eoanthropus is the greater. It will [535] also be noted that there is a marked irregularity in its contour, and that there is a noticeable degree of asymmetry in its conformation, the right side being smaller and not quite of the same form as the left side.
When he had fitted the fragments of the skull into their approximate position, Sir A. Smith Woodward was able to obtain a cast of the cavity which held the brain. So closely does the brain fill the cranial cavity that such a cast reveals not only the size and form of the brain, but also many of the finer markings which give the anatomist a clue to the actual anatomy of the brain. The brain or endocranial cast was entrusted to Professor Elliot Smith for examination. His verdict, pronounced after his first examination, was that, "taking all its features into consideration, we must regard this as being the most primitive and most simian human brain so far recorded."
When we sum up all the characters which Sir A. Smith Woodward has portrayed in this new form of being–the anthropoid characters of the mouth, teeth, and face, the massive and ill-filled skull, the simian characters of the brain and its primitive and pre-human general appearance –one feels convinced that he was absolutely justified in creating a new genus of the family Hominidæ for its reception. This new genus he named Eoanthropus– dawn-man. Ever since Darwin impressed the truth of his theory of man's origin on his fellow-scientists we have expected to encounter man's progenitors, but no one, so far as I know, ever anticipated the discovery of one showing the remarkable mixture of simian and human characters–such a one as Mr Dawson brought to light at Piltdown.
In such a form was this representative of "dawning" humanity raised from his obscure bed at Piltdown and exposed to the merciless eyes of the scientific world. No sooner had he appeared than he became the subject of debate and the cause of hot contention. Some of his critics, we shall see, still wish to take him to pieces and give his skull to a man of the modern kind and his teeth and jaw to a chimpanzee. Nevertheless he has finally [536] emerged from his ordeal, chastened, perhaps, in many of the ape-like qualities originally attributed to him, as a real human being, but one in whom there still can be detected gross traces of man's simian ancestry. The being who could fashion the Piltdown bone implement was a long way in advance of "dawn-man"; this title belongs of right to the man of Java–Pithecanthropus.
______________________________________________________________________________
1 See Proc. Internat. Med. Congress, London, 1913 (Pathological Section).
2 A later investigation of a larger series of English skulls has shown me that 26.6 cm.2 is an overestimate. As measured in English male skulls the mean palatal area is 25.00 cm.2
3 See p. 215.
4 For details concerning this line or plane–the subcerebral plane–see an article by the author in Journ. of Anat. and Physiol., 1910, vol. xliv. p. 251.
[537] Chapter XXIX: The Difficulties of Reconstruction ...
[558] Chapter XXX: Can Fossil Fragments Yield Reliable Evidence
of Man's Evolutionary History? ...
[578] Chapter XXXI: Heads–Ancient and Modern–in Profile...
[598] So far, we have confined our attention in a general survey of the Piltdown head in profile and satisfied ourselves that we are dealing with one which falls easily, [597] so far as dimensions are concerned, within the modern standard. To appreciate its peculiar features we must go farther afield and see how it compares with other ancient types, such as those found at Galley Hill, at La Chapelle-aux-Saints, in Rhodesia, and at Trinil in Java.
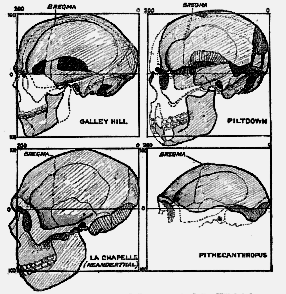
Fig. 217.–Four types of human skulls compared–Galley Hill, Piltdown, La Chapelle-aux-
Saints, and Pithecanthropus.
In fig. 217 are set side by side, within squares of exactly the same size, three of the skulls just named, with the skull of Eoanthropus for comparison. The skull from Galley Hill we may dismiss with a brief note. It represents the modern type of man, and from the analysis just given it will be seen to have many points in common with the skull of [598] Eoanthropus....
[603] Chapter XXXII: The Brain of Fossil Man ...
[637] Chapter XXXIII: The Piltdown Mandible...
[648] If the reader will place a finger in front of the opening of the ear he will feel the condyle
enter its cavity as the lower teeth close against the upper. The position of the various parts of the jaw, when the mouth is opened, is also shown in fig. 237. A rotatory movement of the ascending ramus occurs as the mouth is opened. The condyle then mounts the articular eminence. A strong muscle, the external pterygoid, drags the condyle forwards on the eminence, thus depressing the body of the jaw and opening the mouth. It is a general law of the animal body–one established a hundred and fifty years ago by John Hunter–that no muscle can act by itself; its opponents–the muscles which produce an opposite movement–must act at the same time to a moderate and yielding degree. The external pterygoid has three strong opponents–muscles which close the lower jaw and teeth against the upper jaw and teeth, thus masticating the food. One of these is the temporal muscle, which rises from the side of the skull and is attached to the[649] coronid process (fig. 237). The two others are also important for our present purposes and must be mentioned. One is the masseter, which rises from the zygoma (fig. 237) and is attached to the outer surface of the ramus of the jaw, especially in the neighbourhood of the angle. On the deep surface of the ramus there is a counterpart to the masseter muscle–the internal pterygoid. Now, it will be seen from our illustrations that as the mouth opens under the action of the external pterygoid and the forward movement of the condyle is initiated, the coronoid process is moved forwards and downwards, elongating the temporal, while the angle moves backwards and downwards, stretching the masseter and internal pterygoids, bringing them into an advantageous position for executing a grinding movement. The part played by the articular eminence in the mechanism of opening the mouth will be evidenct. The resistance it offers to the forward movement of the condyle ensures a rapid opening of the mouth. It does more. When the condyle has mounted the eminence, the lower series of teeth is nearly parallel to the upper, not divergent, as would be the case if the joint movement were of the hinge type. When the three great muscles press the lower teeth against the upper, crushing the food, the condyle slips home to its socket–the glenoid cavity in front of the ear. It will be seen that, as the teeth meet and the movement comes to an end, there must be a certain degree of rubbing between the teeth, for the condyle passes backwards as it sinks home and the condyles do not come to rest exactly at the same point of time, giving the teeth a lateral movement. We expect, therefore, that the chewing surfaces of the Piltdown teeth should be worn flat, for the highly developed articular eminence and deep glenoid cavity proclaim in unmistable terms that the chewing movement just described as true of modern man are also true of him. The chewing surface of the Piltdown molars are worn smooth and flat. This is a strong point in favour of the authenticity of the jaw. ...
[661] Chapter XXXIV: Evidence of the Teeth of Fossil Man...
It is instructive to reproduce the reconstruction of the Piltdown teeth and mandible which were made before the canine tooth was actually found (fig. 242). On these early reconstructions are superimposed drawings of the teeth and mandible of a modern Englishman. It will be seen [662] that in both reconstructions the Piltdown teeth project in front of and also behind the teeth of the modern man.
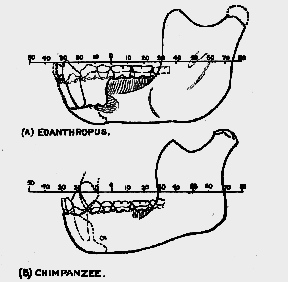
Fig. 242.–A. True profile drawing of the model of the Piltdown mandible and teeth reconstructed under the direction of Sir A. Smith Woodward. Placed on it is the mandible of a modern Englishman. The first molar tooth of the one is placed over the first molar tooth of the other. B. A
reconstruction by the writer. The same modern mandible is used for comparison.
...
[665]
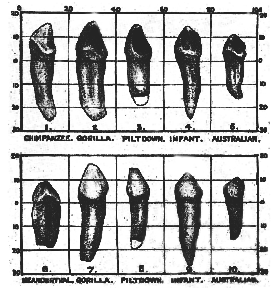
Fig. 243.–The right lower canine tooth found at Piltdown compared with the corresponding tooth of man and of anthropoids.
1. Right lower canine of a young female chimpanzee (lingual aspect).
2. " " " " a female gorilla. "
3. " " " " a child (twice nat. size) "
4. ' ' ' ' an Australian native "
5 Left " " " Neanderthal man "
6 Right " " " a female gorilla (anterior aspect)
7 " " " " Eoanthropus "
8. " " " " an infant "
9. " " " " an Australian native "