On the Discovery of a Palæolithic Human Skull and Mandible
in Flint-Bearing Gravel
Overlying the Wealden (Hastings Beds)
at Piltdown, Fletching (Sussex)
Charles Dawson, F.S.A., F.G.S.
and
Arthur Smith Woodward, LL.D., F.R.S., Secr. G.S.
With an Appendix by Prof. Grafton Elliot Smith, M.A., M.D., F.R.S.
(Read December 18th, 1912).
Quarterly Journal of the Geological Society Vol. 69 March 1913
[117] I. Geology and Flint-implements [C.D.]
[Plates XV-XVII.]
Several years ago I was walking along a farm-road close to Piltdown Common, Fletching (Sussex), when I noticed that the road had been mended with some peculiar brown flints not usual in the district. On enquiry I was astonished to learn that they were dug from a gravel-bed on the farm, and shortly afterwards I visited the place, where two labourers were at work digging the gravel for small repairs to the roads. As this excavation was situated about 4 miles north of the limit where the occurrence of flints overlying the Wealden strata is recorded, I was much interested, and made a close examination of the bed. I asked the workmen if they had found bones or other fossils there. As they did not appear to have noticed anything of the sort, I urged them to preserve anything that they might find. Upon one of my subsequent visits to the pit, one of the men handed to me a small portion of an unusually thick human parietal bone. I immediately made a search, but could find nothing more, nor had the men noticed anything else. The bed is full of tabular pieces of ironstone closely resembling this piece of skull in colour and thickness; and, although I made many subsequent searches, I could not hear of any further find nor discover anything in fact, the bed seemed to be quite unfossiliferous.
It was not until some years later, in the autumn of 1911, on a visit to the spot, that I picked up, among the rain-washed spoil-heaps of the gravel-pit, another and larger piece belonging to the frontal region of the same skull, including a portion of the left superciliary ridge. As I had examined a cast of the Heidelberg jaw, it occurred to me that the proportions of this skull were similar to those of that specimen. I accordingly took it to Dr. A. Smith Woodward at the British Museum (Natural History) for comparison and determination. He was immediately impressed with the importance of the discovery, and we decided to employ labour and to make a systematic search among the spoil-heaps and gravel, as soon as the floods had abated; for the gravel-pit is more or less under water during five or six months of the year. We accordingly gave up as much time as we could spare since last spring (1912), and completely turned over and sifted what spoil-[118] material remained. we also dug up and sifted such portions of the gravel as had been left undisturbed by the workmen. 1
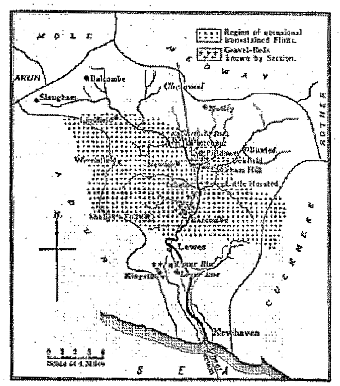
Fig. 1.– Plate of the basin of the Sussex Ouse, showing the distribution of iron-stained flints and flint-bearing gravels.
For many years the harder layers of this gravel-bed have been intermittently worked for farm-road material, as shown by old excavations which are now overgrown, but are traceable over the adjoining fields; and there is known to exist a gravel-bed of appreciable thickness extending over several neighbouring acres. Where the beds have been naturally denuded, a large number of [119] the brown and red flints and the ironstone gravel are to be seen overlying the plough-lands in all directions.
At Piltdown the gravel-bed occurs beneath a few inches of the surface soil, and varies in thickness from 3 to 5 feet; it is deposited upon an uneven bottom, consisting of hard yellow sandstone of the Tunbridge Wells Sands (Hastings Beds). It is composed for the most part of dark-brown Wealden ironstone pebbles, but is mixed, to the extent of about a sixth of the mass, with angular brown flints, a large proportion of which are tabular in form.
2 Occasional cherts and quartzite pebbles also occur, but there are no recognizable Eocene pebbles. 3 The flints vary form 6 or 7 inches in length by 3 or 4 inches in width, down to a very fine gravel or sand. Portions of the bed are rather finely stratified, and the materials are usually cemented together by iron oxide, so that a pick is often needed to dislodge portions–more especially at one particular horizon near the base. It is in this last-mentioned stratum that all the fossil bones and teeth discovered in situ by us have occurred. The stratum is easily distinguished in the appended photograph (Pl. XV) by being of the darkest shade and just above the bed-rock.The gravel is situated on a well-defined plateau of large area, lying above the 100-foot contour-line, averaging about 120 feet at Piltdown, and lies about 80 feet above the level of the main stream of the Ouse. The river has cut through the plateau, both with its main stream and its principal branch, which is called the Uckfield River.
Speaking generally, the remains of this plateau, of which that at Piltdown is merely a part, can in places be traced along a line drawn through Lindfield, Sheffield Park, Buckham Hill, Uckfield, and Little Horsted and southwards, broadening outwards towards the Chalk Escarpment. In fact, the whole country lying between the base of the Wealden Anticline and the Chalk Escarpment presents the appearance of one large low plateau or former base-level plane dissected by the Ouse and its tributary streams.
Remnants of the flint-gravels and drifts have not been mapped or otherwise recorded before in the Ouse Valley, north of the boundary between the Wealden Clay and the Hastings Beds, which runs immediately south of Isfield.

Fig. 2.–Diagrammatic section of the Weald, showing the low plateau on the south capped by flint-bearing gravels.
![]()
Distribution of Flint on the dissected Plateau.
[The thickness of the gravel is much exaggerated in the above section.]
Up to the latter point they are [120] common, and they have been described by Mantell,
4 Topley, 5 and Dr. Elsden 6; but both Topley and those who have followed him use almost the same words, namely, that 'on entering the Hastings Beds country, flints no longer occur either on the surface, or in the river-gravel.' 7 However, in the Tunbridge Wells Sands (Hastings Beds), out of a thousand fields of which I have a record, that have all been thrice tested by means of boreholes to the depth of 3 feet, fifty fields or more furnish abundant evidence of the presence of these brown and red flints. They are present over the Wadhurst Clay between Isfield and Buckham Hill, but the boundary of this stratum is faulted. I have only found them to occur as small fragments, and seldom on the older Wealden Beds of the higher levels within the Ouse watershed.The field-borings go to show that in the greater part of the area over which the flint is distributed, the gravel seldom occurs within 3 feet of the surface in beds of appreciable thickness; but trial-borings along the slopes reveal considerable deposits of gravel, containing small [121] fragments of iron-stained flint, 6 feet and more beneath the surface.
Considering the amount of material excavated and sifted by us, the specimens discovered were numerically small and localized.
Apparently the whole or greater portion of the human skull had been shattered by the workmen, who had thrown away the pieces unnoticed. Of these we recovered, from the spoil-heaps, as many fragments as possible. In a somewhat deeper depression of the undisturbed gravel I found the right half of a human mandible. So far as I could judge, guiding myself by the position of a tree 3 or 4 yards away, the spot was identical with that upon which the men were at work when the first portion of the cranium was found several years ago. Dr. Woodward also dug up a small portion of the occipital bone of the skull from within a yard of the point where the jaw was discovered, and at precisely the same level. The jaw appeared to have been broken at the symphysis and abraded, perhaps when it lay fixed in the gravel, and before its complete deposition. The fragments of cranium show little or no sign of rolling or other abrasion, save an incision at the back of the parietal, probably caused by a workman's pick.
A small fragment of the skull has been weighed and tested by Mr. S. A. Woodhead, M.Sc., F.I.C., Public Analyst for East Sussex & Hove, and Agricultural Analyst for East Sussex. He reports that the specific gravity of the bone (powdered) is 2.115 (water at 5o C. as standard). No gelatin or organic material is present. There is a large proportion of phosphates (originally present in the bone) and a consideration proportion of iron. Silica is absent.
Besides the human remains, we found two small broken pieces of a molar tooth of a rather early Pliocene type of elephant,
8 also a much-rolled of a molar of Mastodon, portions of two teeth of Hippopotamus, and two molar teeth of a Pleistocene beaver. In the adjacent field to the west, on the surface close to the hedge dividing it from the gravelbed, we found portions of a red deer's antler and the tooth of a Pleistocene horse. These may have been thrown away by the workmen, or may have been turned up by a plough which traversed the upper strata of the continuation of this gravel-bed. Among the fragments of bone found in the spoil-heaps occurred part of a deer's metatarsal, split longitudinally. This bone bears upon its surface certain small cuts and scratches, which appear to have been made by man. All the specimens are highly mineralized with iron oxide.[122] Flint Implements.
The brown flints appear to be in every way similar to the plateau flints of Ightham (Kent), and generally to those of the High-Plateau gravels of the North and South Downs. There is the same appearance of tabular and prismatic structure, 'frost fractures,' thick iron-stained patina, and often the same tendency to assume the well-known 'Eolithic' forms (Pl. CVII). There also occur more occasionally certain brilliantly-coloured iron-red flints, presumably more highly oxidized than the prevailing flints, which are of a brown colour. Among the flints we found several undoubted flint-implements, 9 besides numerous 'Eoliths.' The workmanship of the former (Pl. XVI) is similar to that of the Chellean or pre-Chellean stage 10; but in the majority of the Piltdown specimens the work appears chiefly on one face of the implements. They have been very slightly rolled or worn, and, although iron-stained, their patina is not so strong and thick as that of the other flints in the bed. Their form is thick, and the flaking is broad and sparing, the original 'bark,' or surface, of the flint frequently remaining at the grasp, the whole implement thus having a very rude and massive form. Some of them were found on the surface of a ploughed field adjacent to the gravel-bed, which is also strewn with brown and red flints; but, on the surface of the neighbouring plough-lands there occur, in places, flint-implements of various ages, all more or less stained with the iron which abounds in this part of the Weald. In our plates we have confined ourselves to the representation of flints found in the gravel-pit at Piltdown.
As to the 'Eoliths,'
11 it is necessary to speak of them with due reserve and caution. The commonest types belong to the 'borer' and 'hollow-scraper' forms. They occur both in the gravel-bed and on the surface of the plough-lands, and are found in both a rolled and an unrolled (or very slightly rolled) condition. Pl. XVII [123] illustrates both of these classes of 'Eoliths.' Whether natural or artificial, the fractures appear to have been largely governed by the prismatic structure of the flint. Both the rolled and the unrolled 'Eoliths' are deeply stained and patinated, but the former to a much greater extent than the latter.Conclusions.
In conclusion, we may briefly consider the age of the human skull and mandible.
It is clear that this stratified gravel at Piltdown is of Pleistocene age, but that it contains, in its lowest stratum, animal remains derived from some destroyed Pliocene deposit probably situated not far away, and consisting of worn and broken fragments. These were mixed with fragments of early Pleistocene mammalia in a better state of preservation, and both forms were associated with the human skull and mandible, which show no more wear and tear than they might have received in situ. Associated with these animal remains are 'Eoliths,' both in a rolled and an unrolled condition; the former are doubtless derived from an older drift, and the latter in their present form are of the age of the existing deposit. In the same bed, in only a very slightly higher stratum, occurred a flint-implement, the workmanship of which resembles that of implements found at Chelles; and among the spoil-heaps were found others of a similar, though perhaps earlier, stage.
From these facts it appears probable that the skull and mandible cannot safely be described as being of earlier date than the first half of the Pleistocene Epoch. The individual probably lived during a warm cycle in that age.
Explanation of Plates XV-XVII.

G. M. Woodward, del.
Bemrose, Collo, Derby.Plate XV.
Flint-bearing gravel-bed underlying the Tunbridge Wells Sands (Hastings Beds) at Piltdown, Fletching, Sussex. The darkest stratum resting on the bed-rock in the section is that from which the skull and mandible were obtained.
G. M. Woodward, del.
Bemrose, Collo, Derby.Plate XVI.
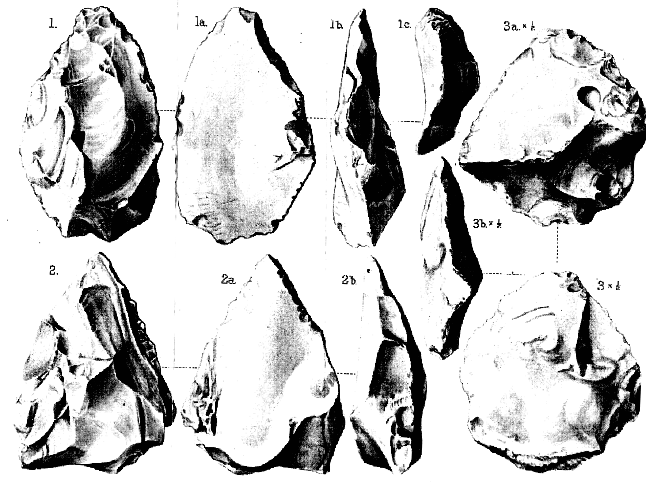
Palæoliths from Piltdown (Sussex)
Fig. 1. Palæolithic implement, well worked on one face (1); a simple flake on the other face (1a );
twisted in edge-view (1 b ); and thickest at the broader end (1c ). Natural size.
2. Palæolithic implement, worked on one face (2); simply flaked on the other face (2 a ); seen
in edge-view (2b ) to be relatively thick at the broader end, with remains of the flint-
module from which it was made. Natural size.
3. Palæolithic implement, showing much of the outer surface of the original flint-module on
its well-flaked face (3); simply flaked on the other face (3a ); and very unsymmetrical in
basal view (3b ). Half of the natural size.
[All the above implements are iron-stained
.]
G. M. Woodward, del. Bemrose.Collo.Derby.
Plate XVII.
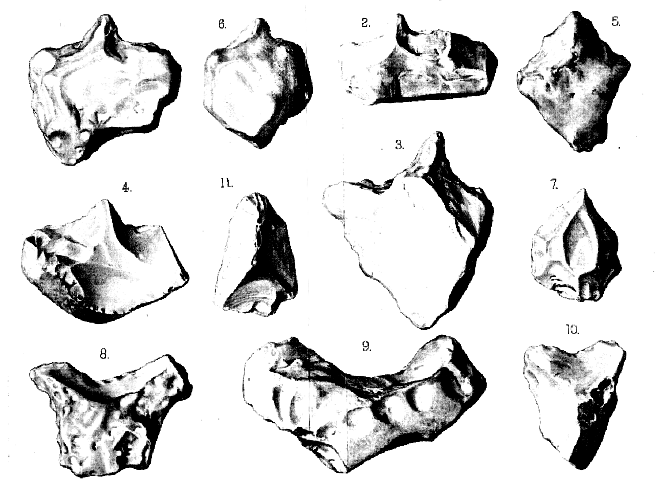
'Eoliths' from Piltdown (Sussex)
Fig. 1-3. 'Eoliths' of the bow-shaped and double-edged scraper type. (Figs. 1 & 3 are
rolled, fig. 2 is unrolled.)
4&5. Similar 'Eoliths' (unrolled) from Fairlight, near Hastings.
6&7. 'Eoliths' of the drill or borer type. (Fig. 6 is rolled, fig. 7 is unrolled.)
8-10. 'Eoliths' of the crescent-shaped scraper type. (Fig. 8 & 9 are rolled, fig. 10 is
unrolled.)
Fig. 11. 'Eolith' (rolled) showing considerable clipping around its point.
[All the figures are of the natural size; and all the flints which they represent
are deeply stained with iron.]
_____________________________________________________________________________
1 These excavations were undertaken with the kind consent of the Lord of the Manor, Mr. G. M. Maryon-Wilson, and Mr. Robert Kenward, tenant of the farm, to whom the Authors wish to record their grateful acknowledgements.
2 There is a tendency among these tabular flints to weather into a prismatic or polyhedral form. One specimen shows a column 2 inches in length. The prismatic structure is well shown in figs. 2, 6, 7, & 9 of Pl. XVII. Most of the modular flints, formerly existing, appear to have disintegrated.
3 Dr. J. V. Elsden writes that Tertiary pebbles are found in the Ouse gravels on the south near Lewes. Eocene pebbles occur in a thin bed east of the Race Stand, on the hill above Lewes, and a similar band is to be seen about 15 feet down the cliff, near the Coastguard Station at Newhaven. See Q.J.G.S., vol. xliii (1887) pp. 616-17.
4 'Geology of the South-East of England' 1833, p. 28.
5 'Geology of the Weald' Mem. Geol. Surv. 1875, pp. 202, 273, 287-88, 292.
6 Q.J.G.S. vol. xliii (1887), p. 646.
7 See also A. J. Jukes-Browne, 'Building of the British Isles' 3rd ed. (1911), p. 426; and A. C. Ramsay, 'Physical Geology, &c.' 5th ed. (1878), p. 344.
8 It is stated by R.A.C. Godwin-Austen (Q.J.G.S. vol. vii, 1851, p. 288) and E. Dixon ('Geology of Sussex' 2nd ed. 1878, p. 110 n.) that remains of the 'large mammalia,' including teeth of elephants, were found in the gravels at Barcombe, 6 or 7 miles south of Piltdown. It is not known to what genera or species these belonged; but, as they are referred to as 'the Asiatic Elephant,' they were probably the true form of Elephas and not Mastodon , though it is possible that some may have belonged to the species akin to E. meridionalis . Specimens from this bed are not known to have been preserved, but they probably belonged to the Pleistocene age.
9 Father P. Teilhard, S.J., who accompanied us on one occasion, discovered one of the implements (Pl. XVI, fig. 2) in situ in the middle stratum of the gravel-bed, also a portion of the tooth of a Pliocene elephant (Pl. XXI, fig. 2) from the lowest bed.
10 Implements of this stage are difficult to classify with certainty, owing to the rudeness of their workmanship. The Piltdown specimens may be compared with an example from Chelles, figured in Piette's 'L'Art pendant l'Age du Renne' p. 36. They resemble certain rude implements occasionally found on the surface of the Chalk Downs near Lewes, which are not iron-stained.
11 The flints, which so nearly resemble those of the plateau gravels of the North Downs, occur sporadically over the South Downs and over many of the older rocks of the Weald. They have lately been discovered in the fields close to the signal-station at Fairlight Down on the summit of the Wealden Anticline (base of the Ashdown Beds and top of the Fairlight Clay) by Mr. W. Ruskin Butterfield, of the Hastings Museum. They occur in the surface-deposits only, mingled with Neolithic implements and certain iron-stained implements which Mr. Reginald Smith, F.S.A., identifies with the early cave types. The last-named are not rolled. Mr. Lewis Abbott, F.G.S., has given considerable attention to this subject, and possesses a large series of implements which we may hope will some day be described in detail.
[124] II. Description of the Human Skull and
Mandible and the Associated Mammalian Remains.
[A.S.W.]
[Plates XVIII-XXI.]
The Human Skull and Mandible.
The human remains comprise the greater part of a brain-case and one ramus of the mandible, with lower molars 1 and 2. All the bones are normal, with no traces of disease, and they have not been distorted during mineralization.
Of the brain-case there are four pieces (reconstructed from nine fragments) sufficiently well preserved to exhibit the shape and natural relations of the frontal, parietal, occipital, and temporal bones, and to justify the reconstruction of some other elements by inference. These bones are particularly noteworthy for their thickness, and for the depth of the branching grooves which are impressed on their cerebral face by the meningeal vessels. The thickening is due to the great development of the finely cancellated diploe, the outer and inner tables of the bone being everywhere comparatively thin. The thickest point is at the internal occipital protuberance, where the measurement is 20 millimetres. A thickness of 11 or 12 mm. is attained at the postero-lateral angle of the left parietal and at the horizontal ridges of the occipital; while a thickness of 10 mm. is observable along the greater part of the fractures of the parietals and frontals. Compared with the corresponding portion on the opposite side, the postero-lateral region of the right parietal is rather thin, its thickness at the lambdoid suture being 8 to 9 mm. It is interesting to add that the average thickness of modern European skulls
1 varies between 5 and 6 mm.; while that of the Australian skulls and of the Mousterian skull from La Chapelle-aux-Saints (France) 2 is from 6 to 8 mm.[125] The largest continuous portion of the brain-case comprises the left side of the frontal and parietal regions, expanding upwards just over the middle line both in the hinder part of the frontal and near the lambdoid suture of the parietal. The position of the middle line is indicated by the impression of the longitudinal sinus on the cerebral face of the bone at both these points, and by a slight longitudinal ridge along the outer face at the hinder end of the parietal region. The left temporal bone is nearly complete and beautifully preserved, lacking only the upper portion of the squamous wing. At one point, it articulates perfectly with the squamous suture of the parietal, and the impressions of the meningeal vessels of the parietal are directly continued on its cerebral face, so that this bone can be assigned to its true position. The left side of the brain-case thus formed has no point of contact with the other two pieces of bone; but one of these is the greater part of the right parietal, extending from the lambdoid suture behind to the anterior end of the squamous suture in front, and so completely symmetrical with the left parietal that its exact position can be determined by measurement. The fourth piece of the brain-case is the middle region of the occipital bone, with a trace of the border of the foramen magnum below and a short extension as far as the lambdoid suture in one part of the right side above. The occipital and the right parietal thus come into direct contact for an extent of about 20 mm. The greater portion of the brain-case may, therefore, be reconstructed without any hypothetical restoration; the only serious deficiency being the middle portion of the frontal region above the supraorbital ridge. Such a reconstruction, with a justifiable amount of modelling, has been skilfully made by Mr. Frank O. Barlow in the Palæontological Laboratory of the British Museum, and this is shown from four aspects, with a longitudinal median section, in Pl. XVIII, figs. 1-4, and text-fig. 3 (p. 126). The actual pieces of bone are indicated by a dark tint, and the modelled portions by a lighter tint, while the hypothetical part is white.
As seen from above (norma verticalis, Pl. XVIII, fig. 1), the cranium is very wide posteriorly, measuring 150 mm. across the widest part just behind the malar arch, and tapering forwards to a slight constriction behind the supraorbital ridge, where its width is 112 mm. The total length from the middle of the supraorbital ridge (glabella) to the external occipital protuberance (inion) is a little uncertain, owing to the hypothetical restoration of the middle of the frontas, but it measures probably 190 mm. The cephalic index may, therefore, be regarded as 78 or 79. Owing to the constriction of the frontal region, the malar arch is visible from above.
In anterior view (norma facialis, Pl. XVIII, fig 2), the relative narrowness of the frontal region is well shown, and the roof is seen to rise to the vertex at the widest part of the skull. In side view (norma lateralis, Pl. XVIII, fig. 3), this upward slope is still better seen, and the steeply-curved frontal contour is especially [126] noteworthy. The external occipital protuberance (inion) seems to form the hindmost point of the cranium, though the portion of the occipital immediately above it is in an almost vertical plane.
In back view (norma occipitalis, Pl. XVIII, fig. 4), the contour of the skull is very remarkable. It is relatively low and wide, and gently arched above, with the sides flattened in their upper half, and the mastoid region either vertical or slightly inclined inwards.

Fig. 3.–Sagittal section of the Piltdown skull (black) superposed on that of a skull from Lewes. Levels (dotted), half of the natural size. E = inion of the Piltdown skull; H = inion of the Lewes skull.
The capacity of the brain-case cannot, of course, be exactly determined; but measurements both by millet-seed and by water show that it must have been at least 1070 c.c., while a consideration of the missing parts suggests that it may have been a little more. It therefore agrees closely with the capacity of the brain-case of the Gibraltar skull, as determined by Prof. Keith,
3 and equals that of some of the lowest skulls of the existing Australians. It is much below that of the Mousterian skulls from Spy 4 and [127] La Chapelle-aux Saints, 5 which have a brain-case larger than that of the average modern civilized man.The principal characters of the brain, so far as they can be observed in a cast of the cranial cavity, are described in an appendix to this paper by Prof. G. Elliot Smith (see p. 145).
A detailed examination of the several bones of the skull is interesting, as proving the typically human character of nearly all the features that they exhibit. The only noteworthy reminiscences of the ape are met with in the upward extension of the temporal fossa, and in the low and broad shape of the occipital region.
The frontal region (Pl. XIX, figs. 1, 1a , & 1b ) is complete on the left side and in its upper middle portion, showing that the frontal eminence is regularly and steeply rounded, and separated from the temporal fossa by a sharp ridge, which extends upwards as far as the coronal suture. This ridge (t. ) approaches nearest to its fellow of the opposite side at its upper end, where it is continued by the curved groove on the parietal, which trends still farther towards the middle line. At its lower end the ridge passes outwards on to the external angular process (e.a.p. ), which is short and directly continuous with the slop of the frontal eminence, not separated from this by any depression. The facette for the malar bone (m.f.) is well preserved; part of the smooth concave roof of the orbit is seen (fig. 1b, orb. ); and the sharp supraorbital border is but slightly abraded. It is, therefore, clear that there cannot have been any prominent or thickened supraorbital ridge, and the missing region above the glabella may be restored on the plan of an ordinary modern human skull. The median frontal (metopic) suture is completely obliterated, so far as the bone is preserved above; but a fractured surface shows that the short fronto-alisphenoid suture (s.f.) was only closed in its deeper half, while the coronal suture is still just visible on the wall of the temporal fossa and is conspicuous on the cranial roof. The coronal suture (cor.) is remarkably complicated, and its tortuous folds are seen to occupy a transversely-elongated shallow depression immediately above the limit of the temporal fossa. The total length of the frontal region along the metopic line, from the glabella to the middle of the coronal suture (bregma), must have been from 120 to 130 mm.; while its maximum width at the external angular processes is 125 mm.
Immediately behind the middle of the coronal suture the parietal region is distinctly flattened; but as it expands backwards the roof soon rises to the broad rounded vertex already mentioned, and there is a faint trace of a longitudinal median ridge near the hinder (lambdoid) border of the bone. The parietal boss or eminence is conspicuous on each side above the hinder end of the squamous suture; and this boss forms the apex of a large flattened triangular area, of which the base-line is at the lambdoid suture. The flattening just mentioned is of the same shape on each [128] side, and the conspicuous lines marking the upper boundaries of the temporal fossa are also clearly symmetrical with reference to the long axis of the skull. The upper line, indicating the upper limit of the temporal fascia or aponeurosis, curves upwards at one point to a distance of 36 mm. from the middle line of the cranial roof; while the lower line, which marks the border of the temporal muscle itself, rises to a maximum height of 32 mm. above the summit of the squamous suture, and curves downwards behind long the antero-inferior edge of the parietal flattening already described; but the lambdoid suture is open, and its outer lateral portion is shown to have been deeply serrated or complicated. The mastoid and squamous suture is open throughout its length, and the squamous portion is as well arched as in a typical modern human skull. The antero-inferior angle of the bone seems to have been almost excluded from articulation with the alisphenoid. The cerebral face, though deeply impressed with the grooves for the meningeal vessels, bears no distinct marks of the Pacchionian bodies. The length of the parietal region along the line of the sagittal suture is 120 mm.; while the total length of its border at the lambdoid suture is about 210 mm.
The occipital bone (Pl. XX, figs. 1, 1a, & 1b ) is remarkable, both for its great width an for the relatively large extent and flatness of its smooth upper squamous portion. The depth of this upper portion, from the lambda to the external occipital protuberance, is 55 mm.; while the total length of the curve from the lambda to the middle of the hinder border of the foramen magnum (opisthion, f.mag.), is only 110 mm. The external occipital protuberance (e.o.p.) is distinctly marked, about twice as wide as deep; while the ridges of the superior (v.c l.) and inferior (l.c.l.) curved lines, the median occipital crest (e.o.c. ), and the other usual irregularities for muscle-attachments are also conspicuous. Above the occipital protuberance may be seen faintly the linea suprema (l.s.). The cerebral face of the bone (fig. 1a ) is interesting as showing the unsymmetrical character of the cerebellum–a condition common to modern man of both low and high degree. The grooved horizontal ridge (si.) on the right side of the vertical median ridge is completely above that on the left side, so that the upper surface of the tentorium over the cerebellum (cb.) on this side would be about 15 mm. above that on the left side. It is also important to observe that the external occipital protuberance is below the upper limit of the tentorium, as in modern man; not raised above it, as in the skull of Mousterian man.
The left temporal bone is excellently preserved, lacking only some of the upper part of the squamous wing (Pl. XIX, figs. 2, 2a , 2b, & 2c ). It is typically human in every detail, and corresponds more closely with the same bone in a comparatively modern human skull from an alluvial deposit near Lewes (Brit. Mus. 7571), than with that in the skull of an existing Melanesian from the [129] Chatham Islands (Brit. Mus. 91.1.20.1). The mastoid portion has no conspicuous extension on the occipital plane, and though its process (mast.) is rather small, this is of the characteristic shape and relations, with a deep impression on its inner side for the origin of the digastric muscle. The supramastoid ridge on the outer face behind the malar arch is an irregularly-rounded boss. The malar arch (mal.) arises in the usual form from the squamous part of the bone, and the glenoid cavity (gl.) is as deep as the deepest observable in modern man, with a transversely -extended hollow at the bottom slightly overhung by the anterior ginglymoid surface. There is no 'spina glenoidalis,' such as is described by Prof. Boule in the Mousterian skull from La Chapelle-aux-Saints.
6 A distinct though small postarticular process (p.a.p.) occurs, separating the upper part of the tympanic from the glenoid cavity; and the long axis of the ovoid opening of the external auditory meatus (e.a.m.) is inclined downwards and backwards as in modern man, not forwards as in the Mousterian man from La Chapelle-aux-Saints. 7 The base of the styloid process (st.) proves it to have been very small. The petrous portion of the bone exhibits all its characteristic features, with no peculiarity worth of remark; and the cerebral face of the mastoid bears the usual broad deep groove for the lateral sinus (si.). The maximum horizontal extent of the temporal bone is 95 mm., and the greatest depth of its squamous wing above the auditory meatus is 57 mm.; while its maximum thickness at the parieto-mastoid suture is no less than 14 mm.The various measurements of the parts of the skull already given, and some others that are also of importance, may be conveniently tabulated as below (p. 130), for comparison with the corresponding measurements of the skulls from Gibraltar and La Chapelle-aux-Saints.
The right mandibular ramus (Pl XX, figs. 2, 2a, 2b, & 2c ) is in the same mineralized condition as the skull, and corresponds sufficiently well in size to be referred to the same specimen without any hesitation. It lacks the articular condyle and the upper part of the bone in advance of the molars; but it is otherwise well preserved, and still exhibits the first two molars in their sockets. Its outer face is sufficiently disintegrated to show the direction of the constituent fibres of the bony tissue. The ascending portion, as in the mandibles from Heidelberg and La Chapelle-aux-Saints, is relatively broad, its width just below the sigmoid notch being 45 mm.; while its depth at the coronoid process (cor.) is about 70 mm. As in the same jaws, its hinder margin makes an angle of 110
o with the inferior margin, its sigmoid notch is comparatively shallow, and the neck of its articular condyle (cd.) must have been short. The bone itself is thin, and its outer face is deeply impressed with irregular hollows for[130]
Skull Pilt- Gibr- Nean- La Cha- Spy I Spy II Austral- Lewes
down altar derthal pelle ians Levels
______________________________________________________________________________
Maximum length 190 190 199 208 200 198 190 182
Maximum width 150 144? 147 156 140 150 126 140
Width at supraorbital ridge 125 ... 122 122 114 117 107 110
Width at construction behind
the supraorbital ridge 112 102 107 109 104 106 92 100
Height, basion to vertex 130 ... ... ... ... ... 124 143
Do. basion to bregma 130 ... ... 131 ... ... 125 140
Do. opisthion to vertex 130 ... ... ... ... ... ...
Do. opisthion to bregma 148 ... ... ... ... ... 149 155
Do. vertex to glabella-
ixion line 90 – 80.5 ... 81 87 85.5 100
Do. bregma to the same 84 – 76.5 – 69 69.7 70 88
Do. vertex to glabella-
lambda line ... 55? 57 ... 51 58 56
Horizontal circumference (by
glabella and inion) 540 – 590 600 580 540 518 520
Cephalic index 78 or 79 80? 73.9 75 70 74.80 66.04 76.9
Bregmatic angle 50o 55o? 44o 45.5o 46o 47o 48.30o
Cranial capacity (in cc.) 1070 1080 1230 1626 1500 1650 1190
Total length of sagittal curve 350-360 – ... 357 370
Length of frontal (metopic)
curve 120-130 – 133 121 105 120? 127 110
Do. of parietal curve 120 – 119 121 118 114
Do. of occipital curve 110 – ... 115 122 ... 117 160
Do. from lambda to inion 55 – 51 ... 59 59 ... 55
Do. from inion to opis-
thion 50 ... ... ... ... ... ... 49
Nearest approach of temporal
fashion in mid-line 36 – ... 65 – 60 29 53
Nearest approach of temporal
muscle to mid-line 46 – ... ... ... ... 38 57
Depth of temporal muscle above
squamous suture 32 – ... 35 ... ... 51 45
Maximum depth of squamous wing
above auditory meatus 57 ... ... ... ... ... 39 48
______________________________________________________________________________
Mandible Piltdown Heidelberg Spy I La Chapelle
______________________________________________________________________________
Height of coronoid 70 66.3 60? 60
Width of ascending ramus
just below signoid notch 45 60 – 45?
Depth at m.2 27 31.8 33 31?
Do. at symphysis – 35 38 37
Thickness of m.1 15 20 14 16
Do. at symphysis – 17.5 15 16
Length of m. 1 11.5 11.6 10 –
Breadth of m. 1 9.5 11.2 10.5 –
Length of m.2 12 12.7 10 –
Breadth of m. 2 10 12 10 –
Mandibular or goniac angle
110o 110o – 110oNote.–Unless otherwise stated, the numerals denote millimetres.
Measurements of the fossi skulls cannot be made with great precision, and there are often discrepancies below the middle of different authors. For the Gibraltar and Australian skulls the figures are chiefly those of Prof. Sollas; for Neandethal those of Dr. Schwalde; for La Chapelle those of Prof. Boule; and for Spy chiefly those of MM. Fraipont and Lohest. Some of the measurements of the Australian skull have been kindly taken by Mr. E. S. Goodrich, F.R.S. The estimates of cranial capacity of the Gibralter and Sky skulls are those of Prof. A. Keith. The measurements of the Heidelberg mandible are by Dr. O.. Schoetensack.[131] the insertion of a powerful masseter muscle (ma.). The horizontal portion, or body of the mandibular ramus, measures only about 27 mm. in depth behind, but must have become a little deeper forwards. External to the first and second molars there is the usual prominent oblique ledge (b.) for one of the origins of the buccinator muscle; but this is the only feature visible on the outer face, a large flake of bone behind the position of the mental foramen having been lost when the anterior part was broken. Seen from within, the ascending portion of the ramus is remarkable for the thickening of its anterior margin, to proved a large surface (t.) for the insertion of the temporal muscle as far downwards as the alveolar border. A strengthening ridge extends downwards and forwards from the articular condyle to the lower part of the sharply-defined inner edge of the temporal surface. Below and behind this the large dental foramen (d.) occurs, though its shape is altered by accidental breaking, and still a little lower there is the slight impress of the mylohyoid groove (m.g.). Near the angle (i.pl.) might be expected a roughness for the insertion of a powerful internal pterygoid muscle; but the bone bears only slight irregularities and it is very little curved inwards. Farther forward the inner face of the ramus is curiously smooth, the origin of the mylohyoid muscle, which forms so conspicuous a ridge in man, being not marked even by a faint line.
The great width of the temporal insertion, the situation of the mylohyoid groove behind rather than in line with the dental foramen, and the complete absence of the mylohyoid ridge are all characters of the mandible in apes, not in man. It is, therefore, very interesting to note that as the ramus curves round to the symphysis (s.) its lower margin exhibits an increasingly wider flattening, which begins beneath the second molar, slopes upwards and outwards, and ends in front of a strongly retreating chin. The inner edge of this flattening is sharply rounded, and at the symphysis itself the inner face of the jaw is so much depressed in its lower part that the bone here has the form of a nearly horizontal plate or flange, closely similar to that found in all the apes. The genio-hyo-glossal and genio-hyoid muscles, in fact, must have had their origin in a deep pit, as in the apes; while the digastric can only have been inserted on the edge of the body flange, instead of extending far over the lower border as in man. The absence of the upper part of the symphysis in the fossil is therefore particularly unfortunate, and there is ample scope for speculation as to the precise shape of the bony chin and the extent of the anterior part of the alveolar border. The fractured end does not even suffice to determine whether or not the part preserved reaches the middle line; it merely shows that the walls of the jaw are thin, and that the inner tissue must have been coarsely cancellous. As, however, the whole of the bone preserved closely resembles that of a young chimpanzee, it seems reasonable to restore the fossil on this model, and make the slope of the bony chin intermediate between that of the adult ape and that of Homo heidelbergensis. [132] If this restoration proves to be correct, the length of the alveolar border in front of the molars is 60 mm., instead of 30 to 40 mm., as in all known human jaws; and it seems difficult to fill this space without assuming that a relatively large canine was present.
That the canine in any case cannot have been very prominent, seems to be proved by the remarkable flatness of the worn surface of the molar teeth (Pl. XX, fig. 2c: m. 1, m. 2). Enamel and dentine have been equally worn down by very free movements in mastication, and such a marked regular flattening has never been observed among apes, though it is occasionally met with in low types of men. Although the cusps have been worn down to the plane of the central area in each tooth, very little dentine is exposed–much less, in fact, than is seen in the similarly worn teeth of apes. Both the first and second molars are noteworthy for their considerable length in proportion to their width, each being provided behind with a large fifth cusp. They are constricted in the ordinary manner at the base of the crown (figs. 2 & 2a, m. 1, 2), and in each tooth the two divergent roots are completely separate to their upper end. They are thus very different from some human teeth with fused roots which are claimed to be of Palæolithic age.
8 The first molar measures 11.5 mm. in length by 9.5 mm. in width; while the second molar is larger by 0.5 mm. in each direction. The third molar, which is situated almost completely on the inner side of the ascending portion of the jaw, is represented only by its well-preserved socket (fig. 2c, m. 3), which shows that its two divergent roots resembled those of the other molars in not being fused together. The anterior root must have been wider than the posterior root, and impressed by a vertical median groove along its hinder face. The posterior root is shown to have been the thicker antero-posteriorly. The tooth must have been relatively large, not less than 11 mm. in length, and inclined a little inwards. The molar teeth, therefore, although distinctly human, are of the most primitive type, and must be regarded as reminiscent of the apes in their narrowness. The first molar may be compared with a detached specimen already known from Taubach, in Saxe-Weimar. 9A restored model of the mandible, skilfully made to fit the skull by Mr. Frank O. Barlow, is shown from the left side and from above and below in text-figs. 4-6 (pp. 133-37). It assumes that the actual fossil extends just to the symphysis, and the result is distinctly striking. The jaw is rather wide, but the nearly straight molar-premolar series of the two sides converge only gradually forwards; while both canines and incisors are, of necessity, large and space.
While the skull, indeed, is essentially human, only approaching a lower grade in certain characters of the brain, in the attachment for the neck, the extent of the temporal muscles,
[133]
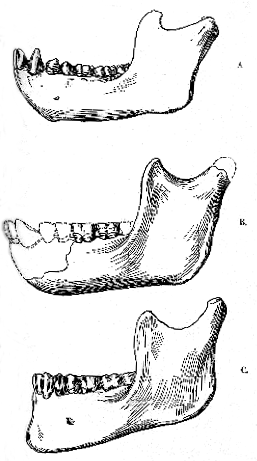
Fig. 4.–Restoration of the Piltdown mandible (B), compared with that of man (C) and the young chimpanzee (A), in left side view; two-thirds of the natural size.
[134]
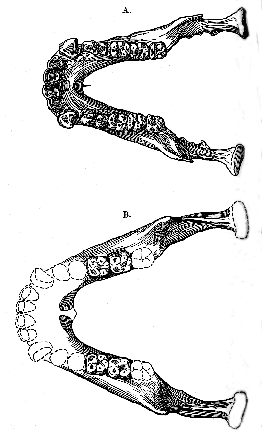
Fig. 5.–Restoration of the Piltdown mandible (B), compared with that of man (C) and the chimpanzee (A), upper view; two-thirds of the natural size.
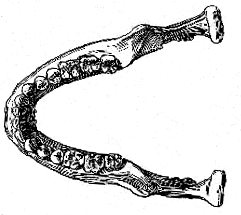
[135] and in the probably large size of the face, the mandible appears to be almost precisely that of an ape, with nothing human except the molar teeth. Even these approach the ape-pattern in their well-developed fifth cusp and elongated shape. The specimen, therefore, represents an annectant type, and the question arises as to whether it shall be referred to a new species of Homo itself, or whether it shall be considered as indicating a hitherto unknown genus.
(Fig. 5 cont.) C.
The brain-case alone, though specifically distinguished from all known human crania of equally low brain-capacity, by the characters of its supraorbital border, and the upward extension of its temporal muscles, could scarcely be removed from the genus Homo; the bone of the mandible so far as preserved, however, is so completely distinct from that of Homo in the shape of the symphysis and the parallelism of the molar-premolar series on the two sides, that the facial parts of the skull almost certainly differed in fundamental characters from those of any typically human skull. I therefore propose that the Piltdown specimen be regarded as the type of a new genus of the family Hominidæ to be named Eoanthropus and defined by its ape-like mandibular symphysis, parallel molar-premolar series, and narrow lower molars which do not decrease in size backwards; to which diagnostic characters may probably be added the steep frontal eminence and slight development of
[136]
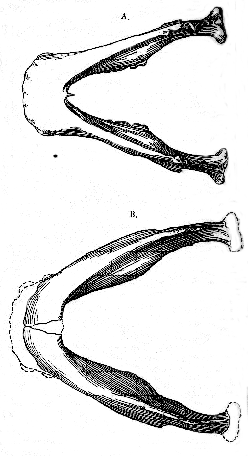
Fig 6. – Restoration of the Piltdown mandible (B), compared with that of man (C) and the chimpanzee (A), lower view; two-thirds of the natural size.
[137] brow-ridges. The species of which the skull and mandible have now been described in detail may be named Eoanthropus dawsoni, in honour of its discoverer.
The difference between the mandible of E. dawsoni and other primitive human mandibles hitherto discovered are very remarkable; but they are especially striking when comparison is made between the new specimen and the mandible of Homo heidelbergensis (fig. 7, p. 138) 10 These two jaws can be very satisfactorily compared; because the first and second molars are almost identical in length in the two specimens, and their outline may be exactly superposed, as shown in fig. 7.
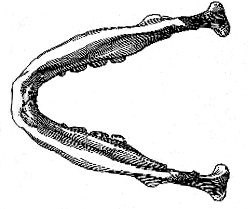
Fig. 6 (cont) C
.
While separated by the characters of the inner face and symphysis already noted (p. 131), the mandible of Eoanthropus dawsoni is thus seen to differ also in the comparative slenderness and its regular deepening to the symphysial end; and, whereas the anterior teeth of the new specimen must have occupied a space of about 60 mm., those of Homo heidelbergensis have only the usual extreme human exent of 40 mm. When it is remembered that Eoanthropus dawsoni and H. heidelbergensis are almost (if not absolutely) of the same geological age, we are thus led to the interesting conclusion that at [138] the end of the Pliocene Epoch the representatives of man in Western Europe were already differentiated into widely divergent groups.
The skull is equally remarkable when compared with the other undoubtedly ancient human skulls hitherto known, and suggests generalizations of even wider import. The discoveries of the brain-case of Pithecanthropus and several skulls of the Mousterian (Neanderthal) type have led to the very general belief that early man was characterized by a low, flattened forehead and a prominent bony brow, like the corresponding parts in the adult existing apes. The only opinions to the contrary have been based on discoveries of very doubtful authenticity, or on theoretical considerations which still need to be tested by more facts. Now, the Piltdown specimen, which is certainly the oldest typically-human brain-case hitherto found, exhibits no anterior flattening, but has the frontal eminence as steep as in modern man, without any prominent supraorbital ridge.
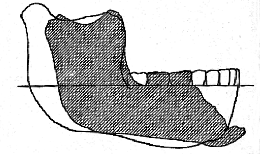
Fig. 7.–Mandibular remus from Piltdown superimposed on that of Homo heidelbergensis; two-thirds of the natural size.
The small development of this ridge may possibly be due in some degree to the circumstance that the new specimen represents a female, as suggested by the small backward extent of the temporal muscles, the weakness of the mandible, and the relative small size of the mastoid processes. Even so, however, a full-grown male of the same race could not have developed a supraorbital prominence approaching that of Mousterian man. The conclusion seems therefore inevitable, that at least one type of man with a high forehead was already in existence in Western Europe long before Mousterian man, with a low and prominent brow, spread widely in this region. It is also clear that this earlier man had a much lower cranial capacity than most examples of the later low-browed man. We are thus reminded of the interesting fact [139] that, during the post-natal life of all the existing apes, the skull has at first the curiously rounded shape of the Piltdown specimen, with a high frontal eminence and scarcely any brow-ridge while as growth proceeds a postorbital constriction begins, the bony brow grows forwards, the forehead becomes flattened, and the familiar well-marked ape-skull is the result. (fig. 8, p. 140). Our knowledge of the principles of palæontology compels us to suppose that the full-grown skull in the ancestral mid-Tertiary apes was of the immature rounded shape just mentioned, although we have not yet been fortunate enough to discover an example; and, during the lapse of Upper Tertiary time, the skull-type in the whole race of apes has gradually undergone changes which are more or less exactly recapitulated in the life-history of each individual recent ape. Hence, it seems reasonable to interpret the Piltdown skull as exhibiting a closer resemblance to the skulls of the truly ancestral mid-Tertiary apes than any fossil human skull hitherto found. If this view be accepted, the Piltdown type (fig. 9, p. 141) has gradually become modified into the later Mousterian type (Fig. 10, p. 141) by a series of changes similar to those passed through by the early apes as they evolved into the typical modern apes, and corresponding with the stages in the development of the skull in an existing ape-individual. It tends to support the theory that Mousterian man was a degenerate offshoot of early man, and probably became extinct; while surviving man may have arisen directly from the primitive source of which the Piltdown skull provides the first discovered evidence.
For much valuable help in studying these human remains I wish especially to thank Mr. W. P. Pycraft, A.L.S., and Mr. Arthur S. Underwood, M.R.C.S.
The Associated Mammalia
The associated mammalia remains are well mineralized with oxide of ion, and, as might be expected in so coarse a gravel, they are all very fragmentary.
Mastodon (Pl. XXV, figs. 1, 1a, & 1b )–A much-rolled specimen is readily identified as the cusp of a molar of Mastodon, of the same type as M. arvernensis. The outer enamel, with the characteristic irregularities, is well preserved, and the waterworn base shows the upper end of the large pulp-cavity. The cusp has three apices closely pressed together, the median one being relatively small and crushed between the others; and the fossil is sufficiently complete to show that it was an isolated eminence on a tooth, not part of a continuous ridge.
Stegodon (Pl. XXI, fig. 2, 2a, 3, & 3a )–Two fragments of a large Proboscidean molar, which have evidently been broken with great force but are scarcely rolled, are referable to a very primitive type of true elephant. One piece (figs. 2 & 2a ), in which the
[140]
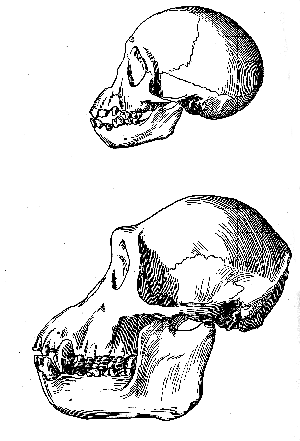
Fig. 8.–Outlines (left lateral view) of the skull and mandible of the young and adult chimpanzee; half of the natural size.
[141]
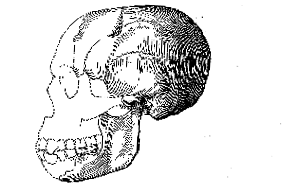
Fig. 9.–Outline (left lateral view) of the skull and mandible of Eoanthropus dawsoni, with the bones of the face and the symphysis of the mandible in dotted outline (1/3 nat. size).
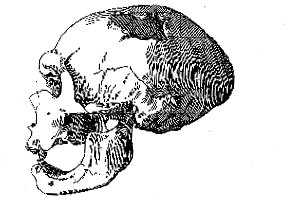
Fig. 10.–Outline (left lateral view) of the human skull and mandible from La Chapelle-aux-Saints, after M. Boule. (1/3 nat. size
[142] ridges are subdivided into round digitations, seems to belong to the hinder part of a molar just coming into wear; while the smaller piece (figs. 3 & 3a ), perhaps of the same tooth, represents the much-worn middle or anterior portion, with the enamel very wavy in section. Both specimens fortunately exhibit a complete valley in cross-section (figs. 2a & 3a ), and so allow the angle of divergence of the ridges to be determined. In the hinder fragment this angle measures about 20
o, in the other a little more; and in each case the walls of the valley are divergent to the summit, not parallel in the upper part. The valleys are thus very wide in proportion to their depth, and the tooth evidently belongs to that primitive section of the genus Elephas to which Falconer gave the name of Stegodon. 11 It cannot be referred to the Upper Pliocene Elephas meridionalis , because in this species the valleys are deeper in proportion to their width, while the ridges are more plate-like and parallel in their upper portion. The new specimen is, therefore, of an earlier Pliocene type, which is best known from the Siwalik Formation in India, and has not hitherto been found in Western Europe. 12Hippopotamus (Pl. XXI, figs. 4, 5, & 5a ).–The broken middle portion of a left molar (probably m. 3) of Hippopotamus (fig. 4) agrees well in size and pattern with many of the corresponding teeth of the ordinary H. amphibius, and may probably be referred to this species. It is only peculiar in exhibiting a small tubercle in the valley on the outer side–an addition which occasionally occurs in more than one species. There is also an example of the first lower premolar (fig. 5 & 5a ), with the usual undivided root. Its crown bears the characteristic fine rugosity, with a trace of the cingulum at the base. Both these teeth are much warn by mastication, and they may have belonged to the same individual.
Cervus elaphus (Pl. XXI, figs. 6 & 6a ).–The base of a large left antler of the common red deer is remarkable for its smoothness, only slight traces of the usual longitudinal furrowing being visible near the burr, which is coarse and prominent. The bone is flattened antero-posteriorly, the hinder face being especially flat, and the plane curving forwards at the origin of the lower brow type. On its anterior face (fig. 6) a rounded longitudinal ridge rises near the outer margin upwards to a thickening at the origin of the upper brow-tyne. The diameter of the base of the beam within the burr is about 70 mm., and the height to the origin of the second brow-tyne is 120 mm.
The collection also includes the proximal end of a metatarsal of Cervus split longitudinally (see p. 121).
[143] Equus.–A first or second left upper molar of an aged horse cannot be specifically determined. The tooth is as broad as long, its enamel is scarcely crimped, and the extent of its inner column is slightly less than half of the antero-posterior measurement of the tooth.
Castor fiber (Pl. XXI, figs. 7, 7a, & 8).–A lower molar and a lower fourth premolar of a beaver, referable to the genus Castor , agree exactly with the corresponding teeth of the Pleistocene and existing species, C. fiber. They exhibit no plication of the enamel such as occurs in the upper Pliocene C. plicidens.
13Of these mammalian remains, the fragment of tooth of Mastodon is so much more waterworn than the others that it may be regarded without hesitation as having been derived from some older deposit. The broken pieces of Stegodon must also have been extremely battered by transport, if their fracture is really natural and not due to the carelessness of workmen. It seems likely, therefore, that these typically Pliocene fossils are of earlier date than the deposition of the Piltdown gravel. The teeth of Hippopotamus , however, may be either Upper Pliocene or Pleistocene in age; while the teeth of Castor are most probably Pleistocene, and typical specimens of Cervus elaphus have never hitherto been found below the Pleistocene. Although, therefore, the mammalian remains do not determine the age of the gravel with certainty, they tend to assign it to the Pleistocene Epoch; and, when it is remembered that (as pointed out by Mr. Dawson, p. 123) most of the contained 'Eoliths' are waterworn, while the associated rude Palæolithic implements have sharp edges, it seems more reasonable to date the deposit by the latter than by the former. If, as the result of these considerations, the gravel proves to have been formed in an early part of the Pleistocene Epoch, the age of the human remains is also decided; for they are very little waterworn, and the skull and mandible would not have occured in close association if they had been transported from the spot at which they were originally entombed.
EXPLANATION OF PLATES XVIII-XXI.
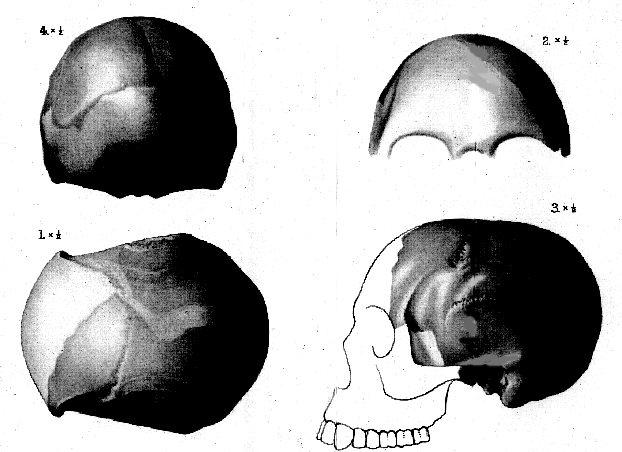
Plate XVIII.
Eoanthropus dawsoni, gen. et sp. nov.; from a gravel near Piltdown Common, Fletching (Sussex).
Fig. 1. Restored model of skull, upper view.
2. Do. do. do., front view.
3. Do. do. do., left side view.
4. Do. do. do., back view.
[All the figures are of half the natural size, and the darkly-shaded parts indicate the pieces of bone actually preserved.]
[144]
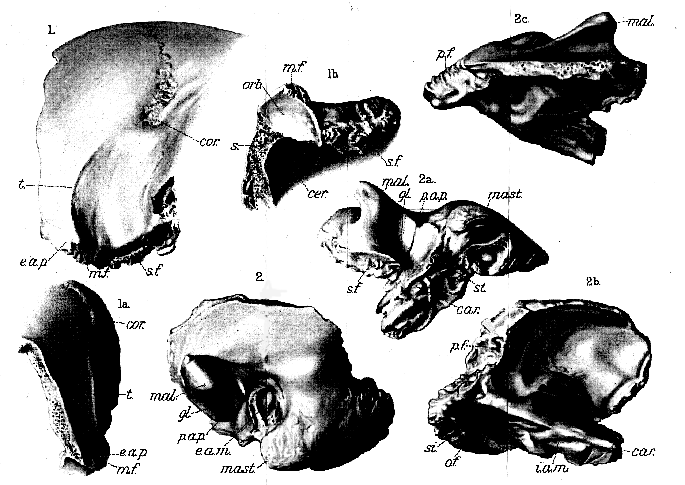
Plate XIX.
Eoanthropus dawsoni, gen. et sp. nov.; from a gravel near Piltdown Common, Fletching (Sussex).
Fig. 1. Left frontal in outer side view; (1a ) front view and (1b ) lower view of the lateral portion.
cerv. = cerebral surface; cor. = coronal suture; e.a.p. = external angular process; m.f. = facette for malar; orb. = roof of orbit; e. = broken section of frontal bone; s.f. = facette for sphenoid; t. = temporal ridge.
2. Left temporal, lacking the upper part of the squamous wing, in outer view; (2a ) lower view; (2b ) inner view; and (2c ) upper view.
car. = openings of carotid canal, the lower blocked by a pebble; c.a.m. = external auditory meatus; i.a.m. = internal auditory meatus; mal. = process for malar; mast. = mastoid process; o.f. = suture for occipital; p.a.p. = post articular process; p.f. = suture for parietal; s.f. = suture for
sphenoid; si. = broad groove for lateral sinus; st. = base of styloid process.
[All the figures are of natural size.]
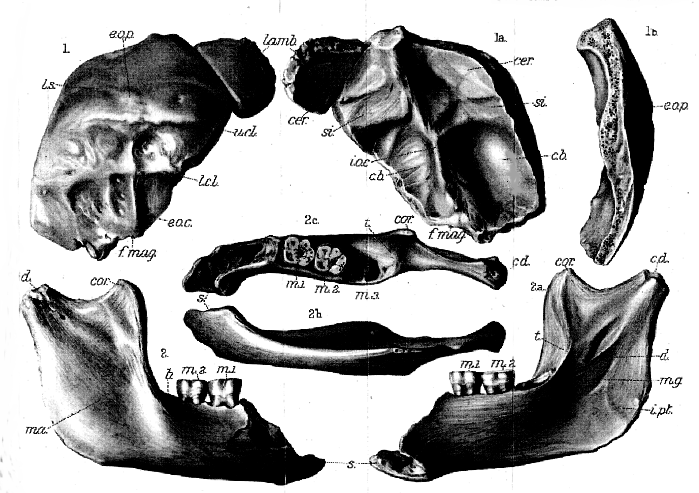
Plate XX.
Eoanthropus dawsoni, gen. et sp. nov.; from a gravel near Piltdown Common, Fletching (Sussex).
Fig. 1. Imperfect occipital in outer view; (1a ) inner view and (1b ) broken vertical section, left side.
cb. = cerebellar fossa; cer.= cerebral fossa; e.o.c.= external occipital crest; e.o.p.= external occipital protuberance; f.mag = foramen magnum; i.o.c. = internal occipital crest; lamb. = portion of lambdoid suture; lc.l = lower curved line; ls. = linea supreme; si. = groove for lateral sinus; u.c.l. = upper curved line.
2. Right mandibular ramus, imperfect at the symphysis, in outer view; (2a ) inner view, (2b ) lower view, and (2 ) upper view.
b. = ridge below origin of buccinator muscle; cd. = neck of condyle; cor. = coronoid process; d. = inferior dental foremen; i.pt. = area of insertion of internal pterygoid muscle; m. 1 ,m. 2 = first and second molars; m. 3 = socket for third molar; m.g. = mylohyoid groove; ma. = area of insertion of masseter muscle; s. = incurved bony flange of symphysis; t. = area of insertion of temporal muscle.
[All the figures
are of the natural size.]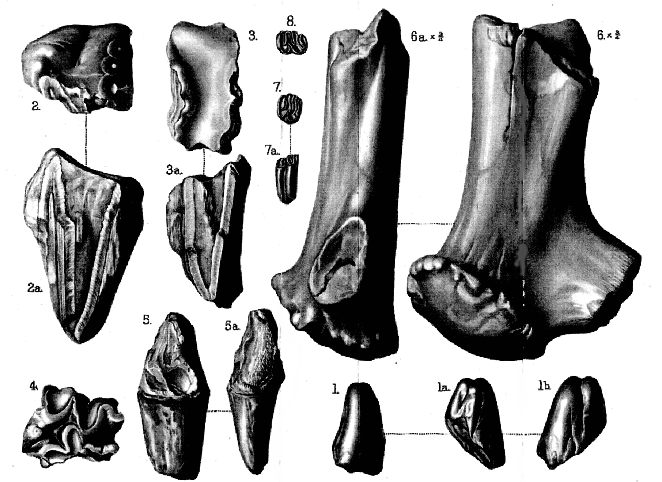
Plate XXI
Mammalian remains found with Eoanthropus dawsoni , in a gravel near Piltdown Common, Fletching, Sussex.
Fig. 1. Mastodon sp.: waterworn cusp of molar tooth, side view; (1a ) front view and (1b ) back view. (See p. 139.)
2. Stegodon sp.: hinder fragment of molar tooth, crown view; and (2a ) vertical longitudinal section. (See p. 139.)
3. Stegodon sp.: fragment of molar tooth worn by mastication, crown view; and (3a ) vertical longitudinal section. (See p. 142.)
4. Hippopotamus sp.: crown view of portion of lower molar tooth. (See p. 142.)
5. Hippopotamus sp.: lower first premolar, outer view; and (5a ) end view.
6. Cervus elephas Linn.: basal fragment of left antler, front view; and (6a ) outer view,
three-quarters of the natural size. (See p. 142.)
7. Castor fiber: first or second right lower molar, crown view; and (7a ) outer view.
(See p. 143.)
8. Castor fiber : right lower fourth premolar, crown view.
[All the figures are of the natural size, except figs. 6 & 6a .]
_____________________________________________________________________________
1. W. J. Sollas, Phil. Trans. Roy. Soc. vol. cxcix n (1908) p. 319.
2. M. Boule, 'L'Homme Fossile de La Chapelle-aux-Saints' Ann. Paleont. vol. vi (1911) p. 20.
3. A. Keith, 'Ancient Types of man' 1911, p. 125.
4. Id. ibid. p. 112.
5. M. Boule, L'Anthropologie vol. xx (1909) p. 261.
6. Ann. de Paleont. vol. vi (1911) p. 58.
7. Ibid. pp. 44, 54.
8. Homo neanderthalensis var. krapinensis, Kramberger, Mitth. Anthrop. Gesellsch. Wien, 1902, p. 191. Homo breladensis, Keith & Knowles Journ. Anat. & Physiol. vol. xlvi (1911), p. 12.
9 A. Nehring, Zeitschr. fur Ethnologie, 1895, p. 338.
10 O. Schoetensack, ‘Der Unterkiefer des Homo heidelbergensis’ Leipzig, 1908.
11 Q.J.G.S., vol. xxi (1865), p. 258.
12 A form of Proboscidean tooth nearly as primitive as that from Piltdown has been recorded from the Pliocene of Lower Austria; see G. Schlesinger, Monatsbl. Verein. Landeskund. Niederosterr, 1911, No. 16.
13 C. I. Forsyth-Major, Proc. Zool. Soc. 1908, pp. 630-31, text-fig. 132.
[145] Appendix
Preliminary Report on the Cranial Cast.
By Grafton Elliot Smith, M.A, M.D., F.R.S., Professor of Anatomy
in the Victoria University of Manchester.
The obserations recorded in these notes were made upon the cranial cast which was submtited to me for examination by Dr. Smith Woodward, under whose direction the remains of the cranium were fitted together and the cast obtained from it.
The accompanying sketch (fig. 11) represents the left norma interalis, and will explain better than a description the extent of missing cranial wall. The greater part of the right parietal bone and a considerable part of the occipital were found, and it was possible to fit them into position. Thus the form and proportions of the whole brain can be estimated.
Fig. 11
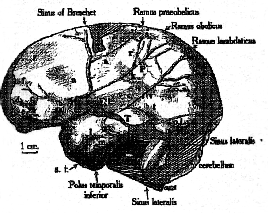
Fig. 11.–Left norma lateralis of the internal cast of the skull from Piltdown.
The sharply-cut meningeal grooves upon the bones have produced upon the cast the whole pattern of the middle meningeal veins and arteries with great distinctness. The diagram shows the arrangement upon the left side: upon the right it conforms to Giuffrida-Ruggeri’s Typus 1. 1
Unfortunately, there are only very slight indications of the [146] arrangement of the furrows upon the surface of the cerebral hemisphere. Nevertheless many of them can be detected, if not by
sight, by passing the finger over the surface and locating the depressions by touch. These features are represented (with considerable exaggeration so far as depth of shading is concerned) in the diagram (fig. 11) on the preceding page.
At first sight, the brain presents a considerable resemblance to the well-known Palaeolithic brain-casts, and especially to those obtained from the Gibraltar and La Quina remains, which are supposed to be women’s. Like these casts this one is relatively long, narrow, and especially flat; but it is smaller, and presents more primitive features than any known human brain or cranial cast.
The position and relations of the cerebellum present a marked contrast with those exhibited in the La Chapelle-aux-Saints, La Quina, and Gibraltar casts; the frontal and parietal regions are less expanded, and the temporal region exhibits some important and very significant differences.
The length of the left cerebral hemisphere (which was probably a little longer than the right, because the left occipital pole was the more prominent, a condition found in the brains of modern right-handed men) 2 is 163 mm. The breadth in the frontal region (just below the point marked a ) is 101 mm.: the maximum breadth of the hemisphere is at the point T, low down on the temporal area, and measures 130 mm.: but at the point P in the parietal region, corresponding to the place where anthropometrists measure the breadth of the skull, the breadth of the brain-cast is only 102 mm. The maximum height, measured from the [restored] tip of the curiously pendant temporal region (denignated ‘Polus temporalis inferior,’ to distinguish it from the temporal pole of the man’s brain) is 100 mm.; but farther back (opposite T) the height sinks to 80 mm.
In this note I do not propose to discuss the significance of the faint glimmerings which this cast affords of the pattern of the convolutions, except to remark that there are indications sufficiently definite to enable us to plot out a great part of the singularly primitive arrangment of sulci.
I have already referred to the diminution and flattening of the frontal and parietal regions. In the centre of the latter there is an area, which is well circumscribed by recognizable sulci (s, n, k )
v, and oa ), raised up into a low hillock, the summit of which is at the point marked P. It is more pronounced on the right hemisphere. This indication of the expansion of an area, the large dimensions and fulness of which are especially characteristic of the human brain, is peculiarly significant, when taken in conjunction with a similar condition of the parietal region.
One of the most striking features of this brain-cast is the deep[147] excavation of the temporal area, to form the wide bay between the inferior temporal pole and the cerebellum. This is due to the marked attenuation of the temporal region; but, as we have already seen in the case of the parietal region, so also here are definite signs tht the expansion has begun which eventually will transform this area into the very different configuration that it presents in the modern brain. There is a very prominent elliptical swelling, the summing of which (at T) is raised more than a centimetre above the level of the surrounding cortex. It is 2 centimetres in vertical measurement and almost 3 centimetres long. This peculiar conformation assumes quite a special interest when it is remembered that this obviously expanding area occupies the position where in the modern human brain is developed the territory which recent clinical research leads us to associate with the power of ‘spontaneous elaboration of speech and the ability to recall names’ (Adolf Meyer).
The configuation of the anterior part of the temporal area is also peculiar, though a suggestion of the same kind of form is seen in the Gibraltar brain-cast. Below the point marked l the surface slopes inwards towards the mesial plane, so tht the fulness of the temporal pole of the modern brain is wanting.
There is marke asymmetry of the cerebellum and of the occipital poles of the cerebrum.
Taking all its features into consideration, we must regard this as being the most primitive and most simian human brain so far recorded; one, moreover, such as might reasonably have been expected to be associated in one and the same individual with the mandible which so definitly indicates the zoological rank of its original posessor.
The apparent paradox of the association of a simian jaw with a human brain is not surprising to anyone familiar with recent research upon the evolution of man. In the process of evolving the brain of man from the ape the superficial area of the cerebral cortex must necessarily be tripled; and this expansion was not like the mere growth of a muscle with exercise, but the gradual building-up of the most complex mechanism in existence. The growth of the brain preceded the refinement of the features and of the somatic characters in general.
______________________________________________________________________________
1 ‘Ueber die endocranischen Furchen der Arteria meningea media beim Menchen,’ Zeitschr. f.
Morph & Anthropol. vol. xv (1912) p. 403.
2 ‘On the Asymmetry of the Caudal Poles of the Cerebral Hemispheres & its Relation to the Occipital Bone’ Anat. Anzeiger, vol. xxx (1907) p. 574.
Discussion.
Sir Ray Lankester congratulated the Authors on the very clear and interesting account given by them of their important discovery. Two distinct questions arose in regard to the bones discovered: first, what are the characters of skull and jaw indicated, and, secondly, what is the geological age of the specimens? He had been allowed to examine the jaw and skull some weeks ago, and with Mr. Dawson had visited the locality where they were found. In regard to the zoological characters of the individual indicated by the bones – everything depended on the little projecting piece af the [148] broken front end of the mandible, proving the existence of a long and flattened symphysis as in the chimpanzee. He considered that Dr. Smith Woodward was fully justified in the conclusion which he drew from this portion of the specimen as to the ape-like character of the completed jaw and of its dentition; and, though the restoration of the jaw on this basis undertaken by Dr. Smith Woodward was a very bold step, he considered that it was justified. He considered that this lower jaw presents simian characters distinct from, and more decisive than, those of the Heidelberg jaw. As to the age of the specimens, he did not think that any conclusion could be arrived at; for the human bones might be earlier than the flints and as early as the Mastodon tooth, or later than either. He did not consider it certain that the lower jaw and the skull belonged to the same individual; and hence no convincing argument as to their age could be drawn from their juxtaposition in the gravel. He would prefer not to use the word ‘Eolith’ to describe any flint-implemente, or supposed flint-implement. Those called ‘Eoliths’ by Mr. Dawson resembled (as did many pieces of the flint) the flints of the High Plateau-gravel of Ightham. The coarsely-worked triangular and irregular flints found in the gravel were certainly of a different character, and probably later. But he did not agree to their being termed ‘Chellean’ or "early Chellean.’ Chelles was simply a locality, and there was no definite ‘form’ of flint-implement which had been designated by the word ‘Chellean.’ It was better to describe the forms of flint-implements, without making use of names for them which had no authorized and accepted meaning and might lead to misunderstanding.
Prof. A. Keith regarded the discovery of fossil human remains just announced as by far the most important ever made in England, and of equal, if not of greater consequence than any other discovery yet made, either at home or abroad. He agreed that the reconstruction of the skull had been executed with great skill, the only point in the restoration about which he was not convinced being the chin-region of the mandible and the form of the incisor canine, and premolar teeth. The restoration approached too nearly the characters of the chimpanzee. The very simian characters of the sub-symphysal region of the mandible, the undoubtedly large anterior teeth, the primitive characters of the skull and brain, seemed to him altogether incompatible with the Chellean age assigned by the Authors. In his opinion the skull must be assigned to the same age as the mammalian remains, which were admittedly Pliocene. In the speaker’s opinion, Tertiary man had thus been discovered in Sussex. In coming to this conclusion the speaker was influenced by the fact that the Heidelberg jaw, which was of early Pleistocene date, the symphysial region of the jaw was essentially human in its markings and characters; whereas the same features in the remains just described were simian, and therefore presumably much earlier.
Prof. Boyd Dawkins said that he agreed with the Authors of the paper that the deposit containing the human remains belonged [149] to the Pleistocene age, and that the Pliocene mammalia in it – Mastodon arvernensis and the rest – had been derived from a Pliocene stratum formerly existing in that area. The latter were merely adventitious, and were no proof of the Pliocene age of the stratum. The Palaeolithic implements were, in his belief, of the same age as the human bones. There was no connexion between the faculty of speech and the capacity for making implements, as was urged by the last speaker. The evidence was clear that this discovery revealed a missing link between man and the higher apes, appearing at that stage of the evolution of the higher mammalia in which it may be looked for – in the Pleistocene age. The modern type of man had no place in this age.
He congratulated the Society on having had the clear and lucid statement of the Authors supplemented by the valuable remarks of Prof. Elliot Smith, the highest authority on the human brain.
Dr. Duckworth agreed entirely with the Authors as to the importance of the Piltdown skull, and also as to its general significance. It was justifiable to associate the various fragments as parts of one human skull; and the simultaneous presence of so many simian characters in one and the same specimen was a point of great significance. Almost any one of those characters might be detected singly in human crania of existing types, especially if search were directed to the more lowly of those. Even the mylohyoid ridge was not so constant as Dr. Smith Woodward suggested. But, so far, the search made by the speaker for a flange-like conformation in a human jaw had been quite unsuccessful. This character, even alone, possessed accordingly the great importance attributed to it by Dr. Smith Woodward. On the anatomical side, the Piltdown skull realized largely the antipations of students of human evolution. To fulfil those anticipations completely, the problem of the precise antiquity of the skull required solution. Anatomists would, therefore, await eagerly the conclusions formed by geologists on this aspect of the subject.
Mr. Clement Reid observed that no detailed ‘drift survey’ had yet been made of this particular area, but perhaps the survey of the Sussex coastal plain might thrown light on the age of the deposit at Piltdown. In the coastal plain the Pleistocene deposits fall into three main groups. at the bottom is the erratic deposit of Selsdy, probably contemporaneous with the Chalky Boulder Clay.
Above comes a series of interglacial deposits showing varying climates and varying amounts of submergence, the submergence culminating in the Goodwood raised beach, at 135 feet above the sea, and passing away in the lesser submergence shown by the raised beach of Brighton. Above all these marine and fluviomarine deposits lies the great sheet of Coombe Rock, which shows a recurrence of Arctic conditions, perhaps dry cold. The uppermost Pleistocene deposit is probably of Mousterian date.
The speaker tried to trace these deposits of the coastal plain continously, through the valleys which breach the South Downs, into the Wealden area, but without much success. It seemed. [150] however, that the low plateau of the Weald, on which the Piltdown deposit probably lies, must belong to a period later than that of maximum depression, for otherwise these lowlands of the Weald would be covered by marine deposits, as is the coastal plain. It was impossible to speak with confidence, but the whole of the evidence suggested that the Piltdown deposit and the plateau on which it rests belongs to a base-level plain, which originated about the period of the Brighton raised beach. The deposits are not pre-Glacial or even early Pleistocene – they belong to an epoch long after the first cold period had passed away; but they occur at the very base of the great implement-bearing succession of Palaeolithic deposits int he South-East of England.
Prof. Waterston pointed out that, if the reconstruction of the cranium and mandible were accepted, it was quite clear that the former was human in practically all its essential characters; while the latter with equal clearness resembled, in all its details, the mandible of the chimpanzee. It was, therefore, very difficult to believe that the two specimens could have come from the same individual. One of the temporal bones, including the glenoid fossa, was complete, and Dr. Smith Woodward had pointed out how closely this bone and the fossa resembled the corresponding parts in modern man. It must be borne in mind that the configuration of the glenoid fossa in man was such as to adapt them for articulation with a human jaw, and not with the mandible as found in the chimpanzee; and, if the jaw had formed part of the skull, it was precisely in the temporal bone that one would have anticipated some variation in structure from the present-day condition.
Mr. A. S. Kennard was of opinion that the gravel-spred in which the remains were found shuld be correlated with the High Terrace of the Lower Thames Valley; the height above the present stream-level was practically the same, and the flint-implements were identical. With regard to the Pliocene age of some of the fossils from Sussex, it must be remembered that the High Terrace of the Thames had yielded several characteristic Pliocene forms. The true succession of the Pleistocene had yet to be worked out, and it must be based on palaeontological evidence.
Mr. Reginald Smith remarked that the flint-implements recovered were of mixed character, and the only course was to date the gravel by the latest specimen. Those exhibited were of the Chelles stage, if not earlier; and, to judge from the photographs shown on the screen, there were hand-axes of the St. Acheul stage in an unfolled condition. While it required a developed brain to manufacture such implements, it was suprising to find so simian a jaw in the later part of the rier-gravel period, and the dilemma still remained. It was a misfortune that the Geological Survey had not visited that area of the Weald in recent years, as it would be interesting to know the relation of the present deposit to the Limpsfield gravel with its homogeneous and well-made implements, [151] which would one day be dated with precision. Successive discoveries justified the adoption of the French classification; and it was idle to decry or ignore the types and terminology that made European archaeologists mutually intelligible, and in fact constituted the grammar of prehistory.
Mr. E. T. Newton called attention to the highly-mineralized condition of the specimens, which seemed to point to their being of Pliocene rather than of Pleistocene age.
The President (Dr. A. Strahan) regretted that, owing to the lateness of the hour, it had become necessary to close this interesting discussion, and called upon the Authors to reply to the points that had been raised.
Mr. Dawson thanked the Fellows for their kind reception of his paper and for their interesting discussion.
He was quite prepared, from an anthropological point of view, to accept an earlier date for the origin of the human remains, and Dr. Woodward and he had perhaps erred on the side of caution in placing the date as early Pleistocene. However, the stratigraphical aspect of the occurrence, as at present understood, compelled them to suggest the comparatively later date for the human remains.
The occurrence of certain Pliocene specimens in a considerably rolled condition, while the human remans bore little traces of rolling, suggested a difference as to age, but not to the extent of excluding the possibility of their being coeval. The rolled specimens might have entered the stream farther up the river than the human remains, and thus might have drifted into the hole or pocket, int he river-bed, where they were found, during the same age but in different condition. Then, again, the skull might have been surrounded by some colloid material which preserved it in its passage from some earlier deposit. It must be admitted that any attempt to fix an exact geological date for specimens found in a gravel-bed is fraught with difficulties.
He expressed his thanks to Mr. S. A. Woodhead for his analyses; to Dr. Elgar Willett for his kind assistance in tracing the gravel; and to Mr. Ruskin Butterfield and Mr. A. W. Pigott for the loan of implements found at Fairlight.
In conclusion, Mr. Dawson expressed his intention of offering the specimens as a gift to the Trustees of the British Museum.
Dr. Smith Woodward admitted that the restoration of the symphysial end of the mandible exhibited was a bold experiment, but he failed at present to conceive of any other interpretation of the fossil. Remembering the failur of Mrs. Selenka’s great excavations in Java where Pithecanthropus wad discovered, he did not anticipate certain success in future work at Piltdown, but he hoped to take part in further diggings. He did not think that the differendes between the Heidelberg and the Piltdown mandibles necessarily implie differences of geological age. The swamps and forests of the Weald in early Pleistocene times may have been a refuge for a backward race.