On the Lower Jaw of an Anthropoid Ape
[Dryopithecus]from the Upper Miocene of Lérida (Spain)
Arthur Smith Woodward, LL.D., F.R.S., Pres. G. S.
(Read April 29th, 1914.)
Quarterly Journal of the Geological Society Vol 70 Dec. 1914
[316] [Plate XLIV.]
Remains of the Anthropoid Apes are so rare among fossils, that every new specimen is worthy of special description and discussion. I therefore offer to the Society some account of the fourth known example of the mandible of Dryopithecus fontani, which has been kindly lent to me by Prof. Luis M. Vidal, of Barcelona. It was found by Senor José Colominas near Seo de Urgel, Province of Lérida (Spain), in a deposit usually ascribed to the Upper Miocene; and, as already observed by Prof. Vidal,1 the specimen is of unusual interest, on account of its occurrence in association with the Hipparion fauna. The jaws of the same species previously discovered in France,2 are from Middle Miocene formations; while the only traces of the great Apes of a later date hitherto found in Europe are a few teeth from the Bohnerz of Würtemberg and the well-known femur from the Sands of Eppelsheim (Hesse-Darmstadt).
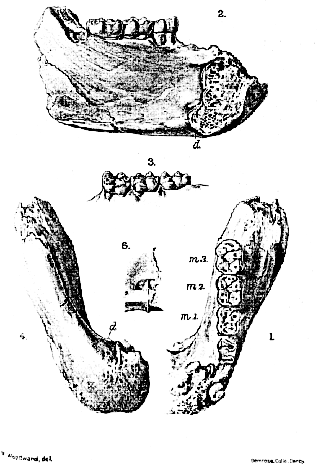
The new specimen (Pl. XLIV) comprises the greater part of the left mandibular ramus and the lower half of the symphysis, with the three molars well preserved and the roots of the fourth premolar broken in the socket. The pattern of the molars and the steepness of the symphysis determine the generic position of the fossil; while the proportions of the bone and the characters of the molar teeth are so closely similar to those of the specimens of D. fontani from the type-locality, that (as already recognized by Prof. Vidal) it can only be referred to this species. The outer cusps of the molars are more worn than the inner cusps in the usual Ape-fashion (Pl. XLIV, fig. 1); and each antero-external cusp (protoconid) is flanked by the short cingulum characteristic of the genus (Pl. XLIV, fig. 3). The enamel is perfectly smooth, and the only crimping that might produce a small secondary cusp occurs between the two inner cusps on the third molar. The first molar [317] (m. 1) measure only 10 mm. in length, and is somewhat narrower than the other, with a relatively prominent antero-internal cusp (metaconid). The second (m. 2) and third (m. 3) molars measure 11 mm. and 11.5 mm. in length respectively, and the third tapers a little behind where the posterior cusp (mesoconid) is relatively large. Of the fourth premolar it can merely be stated that the roots prove it to have been as large as the same tooth previously described in Dryopithecus fontani. [318]
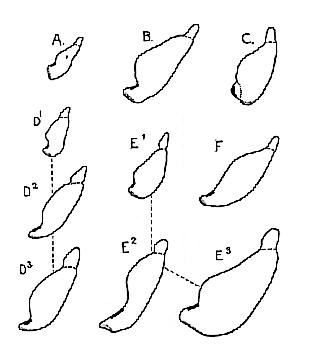
Fig. 1.–Cross-section of the mandibular symphysis in various Primates; two-thirds of the natural size.
A.. Adult Mesopithecus. B. Adult Dryopithecus. C. Adult Homo heidelbergensis. D.1 Very young Chimpanzee, with (D2) young adult and (D3) adult female of the same. E.1 Very young Gorilla, with (E2) adult female and (E3) adult male of the same. F. Adult Eoanthropus dawsoni.. The dotted border marks the insertion of the digastric muscle, and the thick black border the origin of the geniohyoid.]
[318[ The bone is remarkably stout, and the symphysis, so far as preserved, is as heavy as that in the mandible described by Gaudry. As well shown in upper (Pl. XLIV, fig. 1) and lower (Pl. XLIV, fig. 4) views, the right ramus is broken away a little to the right of the median line; and, as seen in inner view (Pl. XLIV, fig. 2), the upper ledge of the symphysis is partly obscured by hard matrix. The inner face of the mandibular ramus is accidentally indented by crushing in the lower part, but the symphysis evidently retains its original shape. The smooth bone bears no clear mark of the floor of the mouth (mylohyoid muscle), but the limits of the insertion of the large digastric muscles are distinct (Pl. XLIV, fig. 4, and text-fig. 1 b, P. 317.) The surface for the digastrics (d.), which is divided in the middle line by a slight ridge, truncates the lower border of the symphysis obliquely, being directed both backwards and downwards; and the vertical thickness of the muscle at this insertion must have been scarcely less than 10 mm., or at least a quarter of the maximum depth of the jaw. From the anterior border of the digastric insertion the plane of the outer face of the symphysis inclines upwards abruptly, without any horizontal extension below (Pl. XLIV, figs. 2 & 4). Above the posterior border of the digastric insertion the inner face of the symphysis is impressed with the usual deep pit (Pl. XLIV, fig. 5), in which the two ovoid hollows for the origin of the geniohyoid muscles are seen. Above this again the inner face of the symphysis rises sharply, but soon turns forward into the long ledge sloping upwards to the front teeth. As shown in the broken section (Pl. XLIV, fig. 2), the cancellous tissue of the bone is of very open texture, and the outer dense wall appears to be unusually thin.
The horizontal ramus of the mandible of Dryopithecus, with its symphysis, is therefore well known, and the two opposing views as to the systematic position of this Ape may now be reconciled in the light of recent researches. In his original description, Lartet considered that the apparent shortness of the jaw implied a face more nearly human than that of the existing Anthropoids, and he pointed to other features which suggested some approximation to Man. Gaudry, on the other hand, after studying a better-preserved specimen, decided that Dryopithecus was the lowest of the known Apes, and even approached the Macaques in the shape of the mandibular symphysis.
If the transverse section of the mandibular symphysis of Dryopithecus (text-fig. 1 b) be compared with that of the nearly contemporaneous Macaque, Mesopithecus (text-fig. 1 a, p. 317), a striking resemblance will indeed be noted. There is the same abrupt upward slope of the anterior face of the bone, and although the digastric insertion (dotted in the figure) is relatively smaller in Dryopithecus than in Mesopithecus, it is still very large for an Anthropoid. The digastric muscles in Macaques and other Monkeys are, in fact, excessively developed to undertake some of those functions of the floor of the mouth that are performed by the mylohyoid and geniohyoid muscles in the large existing
PL. XLIV.
DYROPITHECUS FONTANI Lartet.
[319] Anthropoids. 1 in the latter the digastric muscles are more or less reduced, and Dryopithecus represents the stage in which the reduction had only just begun.
So far as the shape of its mandible is concerned, Dryopithecus is, therefore, a primitive type; and it is interesting to add that the relatively small size of its first molar is a character of the Macaques and other monkeys rather than of Anthropoids, although it is observable in some Gibbons.
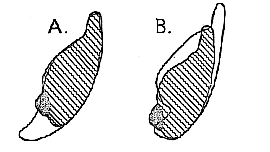
Fig. 2.–Cross section of mandibular symphysis of Homo heidelbergensis (shaded), superimposed (A) on that of Eoanthropus dawsoni (outline) and (B) on that of Dryopithecus fontani (outline); natural size. Secondary bone at the origin of the geniohyoid muscles dotted.
A comparison of the mandibular symphysis of Dryopithecus with that of the existing Chimpanzee, Gorilla, and Orang, shows that it is also of a generalized type. In adult individuals of these Apes, the lower border of the symphysis is extended backwards either into a slender bony flange (text-fig. 1 D2, D3, & E2, p. 317) or into a very massive thickening (E3); consequently, the anterior face of the bone does not slope abruptly upwards from the front border of the digastric insertion, but only turns upwards after a more or less marked horizontal extension. In other words, the bony chin is antero-posteriorly elongated. In newly-born individuals of all these Apes, however, there is no such elongation or extention, and the anterior face of the symphysis (text-fig. 1 D1 E1) slopes upwards almost as abruptly as in Dryopithecus. The Miocene Ape may, therefore, have been a common ancestor of the existing Apes in question.
The generalized character of the mandibular symphysis of Dryopithecus is still further emphasized when comparison is made with that of the oldest known species of Homo (H. heidelbergensis, [320] text-fig. 1 c, p. 317). Here it is necessary to remember that the pit on the inner or posterior face of the symphysis is nearly filled by secondary bone developed in the tendon at the origin of the geniohyoid muscles. The postero-inferior outline to be compared is, therefore, indicated by a dotted line in the figure. Allowance being made for this, the resemblance is seen to be remarkably close–the anterior face in Heidelberg Man being only steeper, and the digastric insertion now facing more directly downwards.
The mandibular symphyses of Mesopithecus, Dryopithecus, and Homo heidelbergensis form, indeed, a gradational series (text fig. 1 A, B, & C), in which there appear to be no place for a stage resembling that of any adult existing Ape. It is difficult even to understand how Eoanthropus (text-fig. 1 F) can be one of the series. If the outlines of Eoanthropus and Homo heidelbergensis be superposed (text-fig. 2 A, p. 319), it will be observed that the former differs from the latter in the specialization of the lower border towards that of a modern Ape. If the outlines of Dryopithecus and Homo heidelbergensis be superposed (text-fig. 2 B), the difference is seen to lie mainly in the reduction of the tooth-bearing region in the latter. On theoretical grounds, the second form of reduction seems the more likely to have taken place.
Hence, so far as the mandibular symphysis is concerned, the Miocene Dryopithecus resembles the large modern Anthropoids no more closely than it agrees with the earliest known true Man. By slight changes in two different directions it may be passed into the one as readily as into the other.
EXPLANATION OF PLATE XLIV.
Dryopithecus fontani Lartet; left mandibular ramus and symphysis, natural size.–Upper Miocene; Seo de Urgel, Lérida (Spain).
Fig. 1. Upper view, showing crowns of molars.
2. Inner view, showing cross-section near symphysis.
3. Outer view of molars, showing cingulum.
4. Lower view showing symphysis and digastric insertion.
5. Inner view of symphysis, showing pits for origin of geniohyoid and genioglossal muscles. d. = insertion of digastric muscles; m. 1-3 = molar teeth.
DISCUSSION
Mr. R. B. Newton said that he was of opinion that the terms Pliocene and Miocene, or those of any of the other great periods, so frequently applied in palæontological research-work, were inadequate for accuracy, and that it was necessary to quote the particular stage of those rocks which had yielded the material described. He instanced the Pikermi Beds, which were often referred to the Lower Pliocene, whereas they belonged to the latest stage of the Miocene, known as the Pontian. To the same part of the Miocene would belong the specimen described by the Author, on account of its association with Hipparion.