I Supplementary Note on the Discovery
of a Palæolithic Human Skull and Mandible of Piltdown (Sussex)
Charles Dawson, F.S.A., F.G.S., and Arthur Smith Woodward, LLD., F.R.S., Pres.G.S. With
an Appendix by Prof. Grafton Elliot Smith, M.A., M.D., V.P.R.S. (Read December 17th, 1913).
The Quarterly Journal of the Geological Society of London Vol. LXX April 1914
[82] [ Plates XIV and XV.]
Contents Page
I. Geology and Flint-Implements 82
II. Description of the Nasal Bones and Lower Canine Tooth of
Eoanthropus dawsoni, and some Associated Mammalian Remains 86
III. (Appendix) On the Exact Determination of the Median Plane of the
Piltdown skull 93
I. Geology and Flint-Implements. [C. D.]
Since reading our paper on December 18th, 1912,
1 we have continued our researches in the Piltdown gravel.The whole bed is divided into four well-defined strata. The topmost consists of surface-soil, containing pieces of iron-stained subangular flint derived from some ancient gravel, similar to those beneath. This surface-soil also contains a mixture of pottery and implements of various ages.
Beneath is the second bed of undisturbed gravel, varying from a few inches to 8 feet in thickness. It is from the centre of this bed that the triangular Palæolithic implement mentioned and figured in our former paper
2 was obtained. It contains rolled and subangular flints similar to those found in the strata above and below, and is mostly pale yellow in colour with occasional darker patches.The third bed, though not always present, is well marked, where it does occur, by reason of its dark ferruginous appearance, and chiefly consists of pieces of ironstone and rolled and subangular flints deeply patinated and iron-stained. As in most other beds of this description, the ferruginous colour of the gravel and of the fossils within it often varies in intensity, from a dull pale-brown to a lustrous blue-black, within the space of a few inches, the latter colour being due to the presence of ferrous sulphide of iron. [83] All fossils found by us (with the exception of the remains of deer) were discovered or have been traced to this third dark bed.
The dark bed rests unevenly upon a fourth, which we identified this year. It consists of very pale-yellow, finely-divided sand and clay, and appears to be a sort of solidified mud reconstructed from the Wealden strata beneath it (Tunbridge Wells Sand, Hastings Beds). This fourth stratum, less than a foot thick, has so far proved unfossiliferous, but contains flints of a much larger size than any of those in the overlying strata. These big buff-coloured flints vary from 6 to 15 inches in length by 3 to 6 inches in width. They are usually flat on one side and uneven and hummocky on the other, which gives them a very characteristic appearance. As they lie in the bed their flat sides are uppermost, and where they impinge upon the bed above they are deeply stained with iron. They are thickly patinated, and saturated to the core with iron.
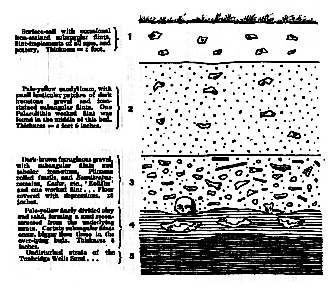
Fig. 1.–Section of gravel-bed at Piltdown (Sussex).
Approximate scale = 1/24th of the natural size.
1 Surface-soil, with occasional iron-stained subangular flints, flint-implements of all ages, and pottery. Thickness - 1 foot.
2 Pale-yellow sandy loam, with small lenticular patches of dark, ironstone-gravel and iron-stained subangular flints. One Palaeolithic worked flint was found in the middle of this bed. Thickness = 2 feet 6 inches.
3 Dark-brown ferruginous gravel, with subangular flints and tabular ironstone. Pliocene rolled fossils, and Eoanthropus remains, Castor, etc., ‘Eoliths’ and one worked flint (Pl. XIV, figs. 1 & 2. Floor covered with depressions. 18 inches.
4 Pale-yellow, finely-divided clay and sand, forming a mud reconstructed from the underlying strata. Certain subangular flints occur, bigger than those in the overlying bed. Thickness = 8 inches.
5 Undisturbed strata of the Tunbridge Wells Sand (Hastings Beds, Wealden).
[84] The character of these flints is often so changed as to resemble ironstone, and some can be scratched and disintegrated with a penknife. This feature may well account for the apparent absence of flints overlying the still higher and older strata in the Weald. They have a roughly-smoothed surface, but bear no striations or 'frost-fractures.' The presence of flints of so large a size, and comparatively little worn and rolled, in the middle of the Wealden area, is a point of great interest. No implements or 'Eoliths' have occurred in this bed, although the latter are fairly common in all the overlying strata.
A few derived chalk fossils have been found by us in the flints from the gravels, and include (according to Mr. C. D. Sherborn and Mr. T. H. Withers):–
Echinocorys vulgaris (= E. scutatus Leske) of the shape characteristic of the zone of Micraster cor-testudinarium.
Inoceramus inconstans (Woods), also a form typical of the same horizon.
The tabular flints so common in the Piltdown gravel are doubtless derived from the regular layers which occur at the base of the zone of Micraster cor-auguinum, and in the upper part of the zone of M. cor-testudinarium. In age, therefore, the derived Chalk fossils and the flints are very similar, belonging to an horizon well exposed at present in the southern Chalk escarpment (as, for example, at Offham) near Lewes.
3Our first work this year was to clear away completely all debris overlying the floor of the dark gravel-bed from the vicinity of the spot where the mandible and piece of occipital bone were found last year, so that the irregularities of its level might be fully exposed. We found that the floor was full of depressions, often measuring 2 to 7 feet across and 1 to 2 feet in depth. Into these depressions had been drifted the dark ferruginous gravel, and in places there yet remained small undisturbed patches. The area so exposed by us measured about 20 feet square.
Following a small rift or channel in the floor which was yet filled with undisturbed gravel we discovered another fragment of a tooth of Stegodon bearing three cusps. This specimen was worked out very carefully, and preserved in the gravel matrix. It seems probable, from its general appearance and condition, that this fragment is a portion of the same molar as that to which the two fragments found last year belonged: like them, it is shattered, and shows little sign of rolling. If so, it must have been broken before its original deposition, and not by the workmen. The other portions were found about 10 yards away, in debris composed of the dark gravel.
In a depression adjoining that in which a portion of the human mandible occurred, was found what appears to be a flint-flake roughly worked on one face and stained dark brown (Pl. XIV, [85]
figs. 1a - [85]1c ); also a triangular flint of Palæolithic outline (Pl. XIV, fig. 2 a-2 c ), but having 'Eolithic' 'edge-chipping' about the apex, the colour and patination resembling those of the 'Eolithic' forms found in the pit generally. Among some of the disturbed gravel in the pit Dr. Smith Woodward found a flint worked on one face and simply flaked on the other face, and similar to the Palæolithic flints described in our last paper.
An incisor of a Beaver (Castor fiber ) and portion of a mandible occurred in the dark gravel: the former close to the spot where the molars were found last year, and the latter some 12 yards away to the south. One of the molars agrees in size with a socket of the alveolar border.
The whole of the work was perforce carried on very slowly, and we found it impossible to employ more shall one labourer, for the actual excavation had to be closely watched, and each spadeful carefully examined. The gravel was then either washed with a sieve, or strewn on specially-prepared ground for the rain to wash it; .after which the layer thus spread was mapped out in squares, and minutely examined section by section.
While our labourer was digging the disturbed gravel within 2 or 3 feet from the spot where the mandible was found, I saw two human nasal bones lying together with the remains of a turbinated bone beneath them in situ. The turbinal, however, was in such bad condition that it fell apart on being touched, and had to be recovered in fragments by the sieve; but it has been pieced together satisfactorily by Mrs. Smith Woodward.
All the gravel in situ excavated within a radius of 5 yards of the spot where the mandible was found, was set apart and searched with especial care, and was finally washed and strewn as before mentioned. It was in this spread that Father Teilhard de Chardin, who worked with us three days last summer, on August 30th, 1913, discovered the canine tooth of Eoanthropus, hereafter described. In this way also Dr. Smith Woodward recovered a small fragment of a tooth of Rhinoceros, in the same state of mineralization as the fragments of teeth of Stegodon and Mastodon..
There now remains little excavation to be done in the immediate vicinity of the site of these remains. Other excavations which we have made in the pit have so far proved unproductive of fossils; but we have opened up some trial-holes which give evidence of a continuation of the bedded gravel to the west, under the ploughland there, and across the small valley on the east near Moon's Farm House.
Conclusions.
We cannot resist the conclusion that the third or 'dark bed' is, in the main, composed of Pliocene drift, probably reconstructed in the Pleistocene Epoch. The evidence seems to point towards a marking-off of the third or 'dark bed' from the second or lighter bed, that contained a triangular Palæolithic implement, [86] one face only of which was worked and was struck in a manner somewhat similar to those of the Chellean stage.
As regards the human cranium discovered including the canine tooth, there is nothing in their mode of occurrence to favour the idea that they may have belonged to different individuals. Putting aside the human remains and those of the beaver, the remains of the fauna all point to a characteristic land fauna of Pliocene age; and, though all are portions of hard teeth, they are rolled and broken.
4 The human remains on the other hand, although of much softer material, are not rolled, and the remains of beaver are in a similar condition. It would, therefore, seem that the occurrence of these two individuals belongs to one of the periods of reconstruction of this gravel, though for other reasons before stated by us, this is not perfectly certain. 5______________________________________________________________________________
1 Q. J. G. S. vol. lxix (1913) pp. 117-44.
2 Ibid. p. 182, footnote 1 & pl. xvi, fig. 2 I find that my former description of these worked flints have been repeatedly misquoted. I took care not to describe these artefacts as of Chellean age; but, for purposes of comparison, I mentioned the work upon one face of the implements as being similar to those of the Chellean stage, that is, one of culture. How far culture can be 'zoned' with age remains to be proved, but there must always exist obvious limitations to such a system.–C.D.
3 'Cretaceous Rocks of Britain: the Upper Chalk' Mem. Geol. Surv. vol. iii (1904) pp. 46-47.
4 That of Stegodon probably presented a more rolled appearance before it was shattered.
5 See Discussion, Q.J.G.S. vol. lxix (1913), p. 151.
II Descriptions of the Nasal Bones and Lower Canine Tooth
ofEoanthropus dawsoni, and some Associated Mammalian Remains. [A.S.W.]
A further study of the cranium of Eoanthropus shows that the slight longitudinal ridge along the outer face of the hinder end of the parietal region is not median, but one of a pair such as frequently occurs in the lower types of human crania. The occipital and right parietal bones thus need slight re-adjustment in the published reconstruction (as described by Prof. G. Elliot Smith in an appendix to this paper, p. 93), but the result does not alter essentially any of the conclusions already reached. The only real addition to our knowledge of the skull is made by the discovery of the nasal bones.
Like the cranial elements, the nasal bones are comparatively stout; and they are thickened at the upper border, suggesting a massive and somewhat overhanging brow-ridge. They are so well preserved that they fit perfectly at the median suture, and show that they form a gently-rounded arch, constricted above and widening below, but with scarcely any upturning at the lower border (Pl. XV, figs. la - 1d ). Both are narrower above than below, and the bone of the right side is considerably wider in its upper part than that of the left side. Below the irregular roughening at the upper thickened end, the outer face of the bone is nearly smooth; while the inner face of each is marked, as usual, with the longitudinal groove for the nasal nerve (Pl. XV, figs. la & ld, n .) The left nasal (figs. 1b & 1c ) is complete, and shows well the extension of the infero-external angle, by which the slightly concave, free lower border of the bone is lengthened. The right nasal (Pl. XV, fig. 1d ) [87] is obviously broken at this angle. The following are some measurements (in millimetres):–
Width of naturally apposed nasals at the upper end 13
Width of naturally apposed nasals at the lower end (about) 15
Length of the median suture 14
Maximum length of the nasals 18
Length of the upper border of the left nasal 7
Do. do. do. right nasal 10
Do. lower do. left nasal 10
Do. do. do. right nasal (about) 11
Comparison proves that these nasal bones resemble those of the existing Melanesian and African races, rather than those of the Eurasian type.
The remains of a turbinal found beneath the nasal bones are too much crushed and too fragmentary for description; but it may be noted that the spongy bone is unusually thick, and has split longitudinally into a series of long and narrow strips.
The remarkable new canine tooth (Pl. XV, figs. 2a -2c ) is certainly that of a Primate Mammal, and may therefore be referred without hesitation to Eoanthropus. As it belongs to the right side of the mandible, corresponds in size with the jaw already found at the same spot, and agrees with the molar teeth in having been considerably worn by mastication, it may almost certainly be regarded as part of the specimen previously described. No trace of the socket for the tooth is seen in the bone preserved at the symphyseal end of the fragmentary mandible, but its position can be determined approximately by reference to the corresponding tooth in the Apes.
The crown of the tooth is conical in shape, but much laterally compressed, so that its inner (lingual) face is concave, while its outer (labial) face is only gently convex. The extreme apex is missing, but whether by wear or by accidental fracture cannot be decided. In the upper half of the outer face (Pl. XV, fig. 2a ) the thin layer of enamel is shown, marked by the usual faint transverse striations (or imbrications); but below this the tooth is encrusted with a film of hydrated oxide of iron, which has broken away at the base of the crown, removing the enamel with it. The darkly-stained dentine is thus exposed here, and the only mark of the lower limit of the crown is a faintly-impressed transverse line just above the constricted neck of the tooth. The enamel on the inner face of the crown (Pl. XV, fig. 2b ) has been completely removed by mastication, while that of the outer face, showing its prismatic structure, is exposed in worn section along the edges of the apical portion. The surface of wear forms a simple gently-curved concavity, evidently produced by a single opposing tooth; and it extends to the basal edge of the crown, as indicated by the clear ending of the cement along its lower margin. It is probably almost parallel with the original inner face of the crown, and the wear has been sufficient near the apex to expose the secondary dentine at the upper end of the pulp-cavity. The
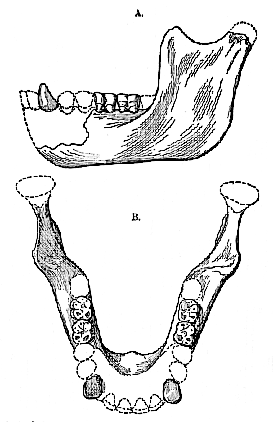
[88] Fig. 2.–Reconstruction of the mandible of Eoanthropus dawsoni in left side view (A) and upper view (B); two-thirds of the natural size.
extent of this pulp-cavity, which is widely open at the lower end of the root, is well shown in two radiographs prepared by Mr. Archibald D. Reid (Pl. XV, figs. 3a & 3b ). It is filled with grains of ironstone or sand. The anterior (median-interstitial) [89] edge of the crown (P1. XV, fig. 2c ) widens below to a triangular tumid area, which is sharply bounded by the outer face, the inner worn face, and the constricted neck of the tooth. Whether or not this area was originally covered to any extent by enamel is uncertain: appearances rather suggest that it is invested, instead of enamel, with a direct combination of the cement-layer of the root. The posterior (lateral interstitial) edge of the crown (P1. XV, fig. 2d ) is sharp, and is not produced below into any inner (lingual) protuberance or heel.
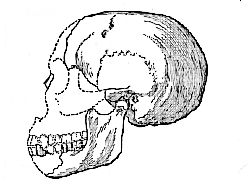
Fig. 3.–Restoration of the skull and mandible of Eoanthropus dawsoni, left lateral view; nearly a third of the natural size.
The root of the tooth is complete, somewhat deeper than the crown, and tapers only slightly downwards from the constricted neck to the truncated lower end. It is irregularly ovoid in transverse section (P1. XV, fig. 2e ), the postero-external (lateral-interstitial+ labial) face being gently convex, while the antero-internal (median-interstitial+lingual) face is flattened. It is invested with a distinct layer of cement, which is seen in cross-section both at the lower end and at the base of the crown on the inner (lingual) face. Much of this cement, especially on the flattened face, appears to be deposited in small, ill-defined, and irregularly-arranged globules.
The following measurements (in millimetres) may be cited:–
Depth of crown preserved (measured along the outer face) 11.5
Maximum (antero-posterior) width of crown 10.0
Do. (interno-external) breadth of crown 10.5
Do. width of the worn inner face 9.0
Depth of root 18.0
Width of flattened face of root at the upper end 9.5
Do. do. do. do. at the lower end 7.0
Smaller diameter of root at the upper end 7.0
Do. do. do. do lower end 5.0
[90] The canine tooth thus described is distinctly larger than any hitherto found in the genus Homo, and differs fundamentally in having completely interlocked with its opposing tooth, which worked downwards on its inner face as far as the edge of the gum. Its exact position in the jaw remains uncertain, but its crown must have risen well above the level of the other teeth, and its state of wear implies its separation from the anterior premolar by a slight diastema, as in the Apes. If the root was originally inclined in a plane almost parallel with that of the anterior end of the jaw, as in the Apes, the crown must have been more nearly upright than that shown in the hypothetical restoration which I made a year ago.
l If, also as in the Apes, the flattened face of the root was almost parallel with the plane of the mandibular symphysis, the concave worn face of the crown would be turned completely backwards and inwards (that is, towards the tongue). In this case (Pl. XV, fig. 4, and figs. 2-3, pp. 88-89) the opposing upper canine tooth must have been shorter and wider than the permanent upper canine in the Recent Apes, for in them the worn surface of the lower canine is continued at least slightly over the outer (labial) side of the crown behind, so that it is visible in internal-external view. There is no evidence of wear by the outer upper incisor, such as sometimes occurs in the Apes; for this wear leaves a small facette in a plane distinct from that made by the canine, also never extends more than about half-way down the crown.The degree of wear of this newly-discovered canine tooth is of especial interest, when considered in connexion with the worn condition of the first and second molars in the mandible to which it apparently belongs. As already described, both these molars are flattened by mastication down to the level of the middle area of their crown, while the third molar (known only by its socket) must have been fully in place (see Pl. XV, figs. 5 a & 5 b ). The permanent canine should therefore be completely extruded and in use, whether the order of appearance of the teeth corresponded with that in Man or with that in the Apes. As, however, the enamel of its inner face is not merely worn, but entirely removed by mastication, the tooth must have been well used for a considerable period. It probably, therefore, came into place before the second and third molars, as in Man–not after one or both of these teeth, as in the Apes.
This appearance of a human order of tooth-succession in the mandible of Eoanthropus suggests the desirability of making a very careful comparison between the shape of the tooth now described, and that of the lower canine in Man as well as in the Apes. As is already known, the permanent canine in some of the Australian and Tasmanian aborigines is comparatively large, with its blunt apex slightly projecting above the level of the dental series; but in this tooth, as in the more normal human [91] permanent canine, the outer (labial) face of the crown is comparatively narrow and deep, and is not continued in a gradual curve round the posterior (lateral interstitial) face to the same extent as in the fossil tooth. The milk-canine of Man, however, may be much more satisfactorily compared. In this tooth the gently-convex outer (labial) face of the acuminate crown (Pl. XV, fig. 6a ) is relatively wider than in the permanent canine, and gradually curves round to the posterior (lateral-interstitial) face, exactly as in the fossil (Pl. XV, figs. 2d & 6d ); while the inner (lingual) face (fig. 6b ) is distinctly concave, and, if its enamel were removed, would correspond very closely with the worn face of the latter specimen (fig. 2b ). Seen from the flattened anterior (median interstitial) face (Pl. XV, figs. 2c & 6e), these two canines appear to be remarkably similar in shape, although, as might be expected, the root is somewhat the shorter in the milk-tooth. In all the existing Apes, and in the extinct Dryopithecus, the permanent lower canine is more conical than that of Eoanthropus , with a more extensive production inwards of the base of the crown on the lingual side. It also appears to be usually larger and stouter in proportion to the size of the jaw. The crown of the milk-canine in the Apes, however, like that in Man, is much more compressed, with a concave inner (lingual) face and a gently-convex outer (labial) face, so that it closely approaches the fossil in shape (Pl. XV, figs. 8a -8d ): the only noteworthy difference being that, in the Apes, there is already the characteristic small produced heel or ledge at the base of the crown, at the hinder (median-interstitial) end of the inner (lingual) face.
It results, therefore, from these comparisons that, among known Upper Tertiary and Recent Anthropoids, the permanent lower canine of Eoanthropus agrees more closely in shape with the milk-canine both of Man and of the Apes than with the corresponding permanent tooth in either of these groups. It is also obvious that the resemblance is greater between Eoanthropus and Homo than between the former and any known genus of Apes. In other words, the permanent tooth of the extinct Eoanthropus is almost identical in shape with the temporary milk-tooth of the existing Homo. Hence it forms another illustration of the well-known law in mammalian palæontology, that the permanent teeth of an ancestral race agree more closely in pattern with the milk-teeth than with the permanent teeth of its modified descendants.
In this connexion, it is interesting to add that even in Homo sapiens, if the base of the crown of the canine were raised in the gum to the same level as that of the adjacent teeth, its apex would frequently project well above the rest of the dental series. The relatively large size and depth in the milk-dentition is especially well seen in a preparation in the Central Hall of the British Museum (Natural History); see Pl. XV, fig. 7.
For valuable help in making these studies, I have again to thank Mr. W. P. Pycraft and Prof. Arthur S. Underwood.
[92] The Associated Mammalia.
Stegodon.
The new fragment of a molar of Stegodon is part of an unworn plate showing three cusps in line, the outermost being smaller and much less elevated than the two others. It evidently belongs to the hinder end of a molar, and it may perhaps be part of the same specimen as the fragments already described.
Rhinoceros (Pl. XIV, figs. 3a & 3b .)
Rhinoceros is represented only by the anterior crest of an upper cheek-tooth, which has been broken and rolled before burial, and is as highly mineralized as the pieces of molars of Stegodon and Mastodon. This specimen has, therefore, the appearance of a derived fossil. As shown in anterior view (fig. 3a ) the crown is rather low, and as seen from above (fig. 3b ) the inner expansion of the crest is very wide at the base. The basal cingulum is preserved only on the anterior face, and does not appear to have extended round the inner end of the crest. The enamel of the inner expansion is almost smooth, but that on the sides of the think part of the crest is marked by a coarse rugosity and a few vertical corrugations. The specimen cannot be specifically determined with certainty; but, on direct comparison, it is found to agree best with the anterior crest of upper premolar 3 of Rhinoceros mercki (= Rh. leptorhinus Owen) and Rh. etruscus . So far as general shape is concerned, it might belong to either of these species; but, as pointed out to me by Prof. Boyd Dawkins, the peculiar coarse rugosity of the enamel is most closely similar to that of Rhinoceros etruscus.
Castor
A fragment of the alveolar portion of the inner face of the right mandibular ramus of Castor corresponds in size with the true molar described last year. 2 This tooth, in fact, fits well the socket for m. 2. The newly discovered lower incisor may also, perhaps, belong to the same mandible.
Explanations of Plates XIV. and XV.
[Ed. note: images reproduced are smaller than those in original plates]
\
Plate XIV.
Fig. 1. Flaked flint, showing few flakings on one face (1a ), a simple flake on the other face (1b), and its tabular shape in edge-view (1c ). Natural size. (See p. 84.)
2. 'Eolith,' showing flaked edge (2a ), simply-flaked face (2b ), and edge-view (2 c ). Two-
thirds of the natural size. (See p. 85.)
3. Rhinoceros cf. etruscus Falconer: imperfect interior crest of right upper premolar 3, front
view (3a ) and inner view (3b ). Natural size. (See p. 92.)
PL. XIV
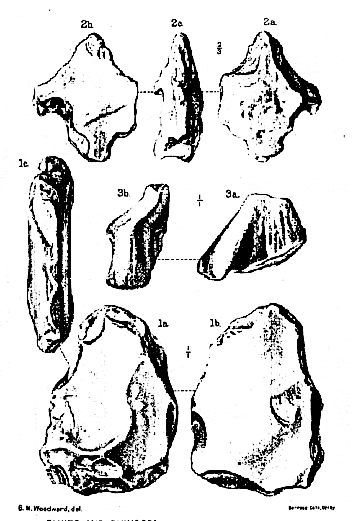
FLINTS AND RHINOCEROS FROM PILTDOWN (SUSSEX).
PL. XV.
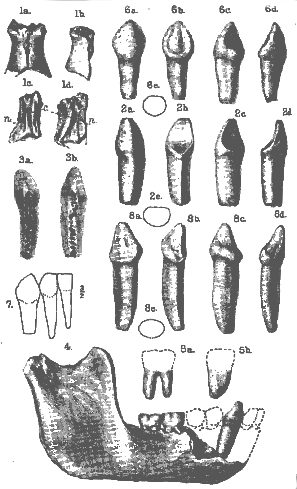
EOANTHROPUS, HOMO, SIMIA.
[93] PLATE XV.
Fig. 1. Eoanthropus dawsoni A. S. Woodward: nasal bones in front view (1a ), left side view (1b ), with left nasal from within (1c ) and right nasal from within (1d ). Natural size. c. = inner crest; n. = groove for nasal nerve. (See p. 86.)
2. Do.: right lower canine tooth in outer or labial view (2a ), inner of lingual view (2b ), anterior or median-interstitial view (2c ), posterior or lateral-interstitial view (2d ), and outline transverse section of neck (2e ). Natural size. (See pp. 87-89.)
3. Do.: radiograph of the right lower canine tooth from without (3a ) and in side view (3b ). Natural size. (See p. 88.)
4. Do.: right mandibular ramus with canine tooth, outer view. Two-thirds of the natural
size.
5. Do.: impression of cavity for roots of lower molar 3; outer (5a ) and anterior (5b ) views, with crown in restored outline. Natural size.
6. Homo sapiens Linn.: right lower milk-canine in outer or labial view (6a ), inner or lingual view (6b ), anterior or median-interstitial view (6c ), posterior or lateral-interstitial view
(6d ), and outline transverse section of neck (6e ). Twice the natural size. (See p. 91.)
7. Do.: right lower milk-canine and milk-incisors, outer antero-lateral view. Three-halves of the natural size. (See p. 91.)
8. Simia satyrus Linn.: right lower milk-canine in outer or labial view (8a ), inner or lingual view (8b ), anterior or median-interstitial view (8c ), posterior or lateral-interstitial view (8d ), and outline transverse section of neck (8e ). Four-thirds of the natural size. (See p. 91.)
_____________________________________________________________________________
1 Q.J.G.S. vol. lxix (1913) p. 133, fig. 4b.
2 Q.J.G.S. vol. lxix (1913) p. 243 & pl. xxi, fig. V.
[93] APPENDIX
.
On the Exact Determination of the Median Plane
of the Piltdown Skull.
Prof. G. Elliot Smith, M.A., M.D., V.P.R.S.
At the meeting of the Geological Society which was held on December 18th, 1912, I gave my first impressions of the cranial cast which Dr. Smith Woodward had sent me three days before the meeting.
On the present occasion it is not my intention to say anything further in reference to the brain of Eoanthropus (because I am preparing a full report upon it for presentation to the Royal Society 1 ); but, as there has been considerable criticism of the restoration of the brain-case, I should like to take this opportunity of expressing my opinion that none of the criticism has affected the accuracy of the preliminary note upon the cranial cast which I communicated to this Society in December 1912. 2
As the correct restoration of the cranium was the necessary preliminary to any detailed study of the form of the brain, Dr. Smith Woodward kindly permitted me to examine the fragments of the skull, and make an independent investigation with the view of determining what positions they originally occupied in the skull. This examination revealed a multitude of structural features which indicate precisely the true position and orientation of each of the fragments; and there is no doubt that the [94] reconstruction of the skull which Dr. Smith Woodward exhibited to the Geological Society in December 1912 was a much closer approximation to the truth than any of the various models so far exhibited in public by his critics.
In the course of my examination of the fragments last November, I found that the anterior end of the sagittal suture was present on the largest fragment (fig. 4, S, below). The recognition of this suture directed my attention to other features of the frontal and parietal bones, which enabled me to determine the precise location (M) of the medial line of the skull.
Fig. 4.–Drawing representing the sutures in the bregmatic region, traced from a photograph; three-halves of the natural size.
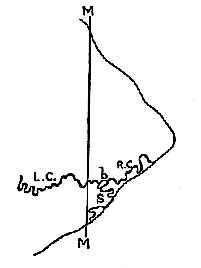
In fig. 5 (p. 95) I have represented diagrammatically the information to be gained from the study of the large fragment when viewed in norma verticalis. The left half of the coronal suture is an obtrusive feature of this fragment. It pursues a very irregular course; but the general direction of the suture is indicated in the diagram by the line L.C. (fig. 5). The exact pattern formed by its medial extremity is shown in fig. 4 (L.C.), which is a tracing from an excellent photograph taken by my assistant, Mr. Henry Gooding.
[95] Careful examination of the fragment also revealed a small part of the right half of the coronal suture (fig. 4, R.C.), the presence of which has already been detected by Dr. Smith Woodward.
Fig. 5.–Diagram of the upper aspect of the largest of the Piltdown fragments.
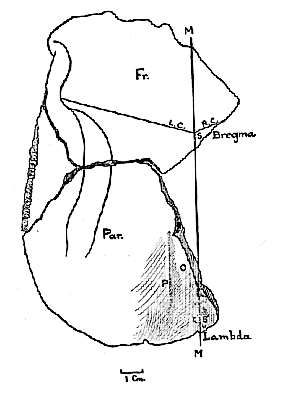
At the place of meeting of the two halves of the coronal suture, which is slightly to the right of the median plane (fig. 4), there is a diminutive bregmatic bone (b). At its posterior border the [96] sagittal suture (S) begins, and pursues a tortuous course towards the broken edge of the bone.
The presence of this part of the sagittal suture affords positive evidence of close proximity to the true median line. The anterior extremity of the sagittal suture often becomes deflected a few millimetres to one side or the other of the median line; but except in those rare cases where a large bregmatic bone is present, the deviation is never considerable. In the course of an examination of the collection of crania now in the Anatomical Department of the University of Manchester, I found several examples in which the anterior end of the sagittal suture closely resembled that of the Piltdown skull (fig. 4, p. 94).
Fig. 6.–Transverse section through the frontal bone, a short distance in front of the bregma.
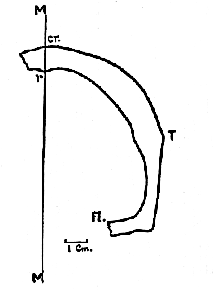
Along the line corresponding to that labelled M in the diagram, a well-defined longitudinal ridge (fig. 6, r, above) was found upon the endocranial aspect of the frontal bone. Careful examination [97] of the fossil shows that this ridge corresponds to the place where the two halves of the frontal bone originally came together at the metopic suture. Although it is not uncommon to find in the crania of primitive man a metopic crest upon the outside of the frontal bone (that is, at the spot marked cr. in fig. 6, p. 96), an endocranial metopic crest is of much rarer occurrence. It is found, however, in the neighborhood of the bregma in the Neanderthal skull; and there is no doubt of its identity in the specimen now under consideration.
Mr. Frank Barlow called my attention to the fact that, if this fragment be looked at from in front (or a transverse section of a cast of it, fig. 6, be studied), the curve of the frontal bone describes a gentle sweep above the temporal ridge (T), which reaches its summit (cr.) directly above the endocranial ridge (r), beyond which it begins its descent, showing that it has crossed the middle line to the right side. This affords independent corroborative evidence of the correctness of the determination of the median plane (M).
My friend Prof. J. T. Wilson F.R.S., has pointed out that the inclination of the floor of the anterior cranial fossa (fig. 6, Fl.) provides further evidence in support of my contention. For, if the bone be tilted laterally to even a small extent, the floor would become inclined at an angle such as is unknown in any skull, whether human or simian.
If the median line thus determined in the region of the bregma (fig. 5, p. 95) is prolonged backwards, it cuts the posterior corner of the parietal in the neighbourhood of the lambda (fig. 5). Independent evidence, derived from (a ) the study of the manner of articulation of the left parietal and temporal bones, and their relationship to the occipital; (b ) the texture of the endocranial surface of the bone along the line M; (c ) the fact that the transverse section of the supralambdoid flattening (O) is horizontal only when the bone is placed as is represented in the diagram; and (d ) the situation and relations of the parasagittal crest (P), – all confirms the accuracy of the identification of the median plane as it is represented in fig. 5.
If the fossil be held in a certain light, a series of depressions (S
1) can be detected, which closely resemble those found in the supralambdoid region of skulls (such as the Neanderthal calvaria) where the sagittal suture has recently become closed. The suggestion is that the posterior end of the sagittal suture conformed to the pattern represented as S1 in fig. 5, and had recently closed when the Piltdown man met his death. This may occur in modern man at any age between 30 and 40 years–although, in rare instances, it may happen before 30, or (more often) be delayed beyond 40. There are reasons for believing that this individual was a young adult, and possibly a female: for the features that present secondary sexual characters in modern skulls are quite indefinite in these fragments._____________________________________________________________________________
1 [Communicated to the Royal Society at the meeting on February 19th, 1914.]
2 Q.J.G.S. vol. lxix (1913) pp. 145-47.
[98] Discussion.
Prof. A. Keith congratulated the Authors on the progress made during the last twelve months.
He was glad to note that the particular stratum in which the remains of Eoanthropus had been
found was being distinguished from the more superficial stratum in which flints of the Chellean type had been found. He ventured to say that, if no human remains had been discovered in the deeper or Eoanthropic stratum, no one would have hesitated in regarding it as of Pliocene age. He was glad to note that the hinder end of the skull of Eoanthropus had been opened out to a considerable degree; but, in his opinion, the occipital and temporal bones were still placed wrongly. When these defects were removed, and the two sides of the skull made approximately symmetrical, it would be found that the brain-capacity was about 1500 c.c. The brain-cast of the skull, as originally reconstructed was just under 1200 c.c.; it was difficult to see how widening out of the skull would reduce the capacity to 1100 c.c. Two other difficulties that he had encountered were (1) the presence of a pointed projecting canine in the jaw, and an articular eminence of the glenoid fossa of the skull; and (2) a much-worn canine tooth in a jaw in which the third molar tooth–according to the published X-ray photograph of the Piltdown mandible–was not completely erupted. He agreed that all these parts–skull, jaw, and canine tooth–must be assigned to Eoanthropus, but he was not convinced that they could all belong to the same individual.
Prof. W. J. Sollas thought that the Authors were to be congratulated on the complete nature of the proof which they had reached by a study of minute anatomical characters. A system of reconstruction which afforded different results from those obtained in this direct and positive manner seemed to require some amendment. The presence of derived fossils in a gravel was one of the commonest facts, not encountered here for the first time, and geologists had followed their usual method in dating the Piltdown gravel by the most recent fossils contained in it; these showed it to be Pleistocene. The topographical relations of a gravel are of some value as evidence, and confirm this conclusion. The precise horizon in the Pleistocene was less definitely known; unfortunately, the flints which had been found in association with the skull were not sufficiently characteristic to determine the point.
Prof. W. Boyd Dawkins said that he would only take up at that late hour, one of the many points raised by Prof. Keith–the age of the Piltdown deposit. It was clearly proved to be later than the Pliocene by the presence of an antler of red deer (Cervus elaphus ), a species unknown in the Pliocene of Europe, and abundant in the Pleistocene and later strata. He agreed with the Authors of the paper in their conclusion that the deposit belongs to an early stage of the Pleistocene Epoch.
[99] Prof. A. S. Underwood said that he would confine his remarks to the two molars and the socket of the third molar in the Piltdown mandible. He had been prepared to show the radiograph at the Royal College of Surgeons in the summer, but Prof. Keith had been unable to place a lantern at his disposal. The two molars were worn down by use, to such an extent that it was impossible that the individual could have been less than 30 years of age, probably a good deal more. The sockets of the third molar were not those of an erupting tooth, the roots had been quite completed, and the tooth was in its final position at death. This was very plainly shown in the radiograph. Had the third molar been erupting or about to erupt, the roots could not have been on a plane with those of the other molars.
Mr. C. Dawson thanked the Fellows for their kind reception of the paper. He specially wished to record the thanks of the Authors to Mr. George M. Maryon-Wilson (the Lord of the Manor) and his tenant, Mr. Robert Kenward, for their kind permission to make excavations in the gravel-bed at Piltdown.
Dr. A. Smith Woodward and Prof. G. Elliot Smith also briefly replied.